CH11.qxd 2/3/05 4:19 PM Page 400
ls
le
ll
spot on it. It pecks at the spot and, within minutes, it has mastered the response needed
to receive a reward.
Now the response requirement can be made more complex. The pigeon might be
required to turn 360° before pecking the disc to gain the reward. The pigeon can learn
this response, too. Other contingencies might then be added, making the response requirements
even more complex. For instance, the pigeon might be trained to turn in a
clockwise circle if the disc is green, to turn in a counterclockwise circle if the disc is red,
and to scratch at the floor if the disc is yellow.
If you suddenly came upon this complex behavior in a pigeon, you would probably
be astounded. But, if you understood the experience that had shaped the bird’s
behavior, you would understand the cause. The rewards offered to the pigeon altered
its behavior so that its responses were controlled by the color of the disc on the
wall.
Skinner extended this type of analysis to include all sorts of behaviors that, at first,
do not appear to be easily explained. For instance, he argued that various phobias could
be accounted for by understanding a person’s reinforcement history. An example is
someone who was once terrified by a turbulent ride on a plane, thereafter avoiding air
travel and manifesting a phobia of flying.
The avoidance of flying is rewarding because it lowers the person’s anxiety level,
and so the phobic behavior is maintained. Skinner also argued against the commonly
held view that much of human behavior is under our own control. From Skinner’s perspective,
free will is only an illusion, because behavior is controlled not by the organism
but rather by the environment through experience.
Although our intent is not to debate the pros and cons of Skinner’s ideas here, we
can conclude that many complex behaviors are learned. It is also true that learning
takes place in a brain that has been selected for evolutionary adaptations. This combination
of learning and inherited brain circuits can lead to some surprising results. A
case in point can be seen, again, in pigeons.
Although a pigeon in a Skinner box can quickly learn to peck a disc to receive a bit
of food, it cannot learn to peck a disc to escape from a mild electric shock to its feet.
Why not? After all, the same simple pecking behavior is being rewarded. But apparently,
the pigeon’s brain is not prewired for this second kind of association. The bird is
prepared to make the first association but not prepared to make the second, which
makes adaptive sense.
For a pigeon, pecking is a behavior that in a natural environment is widely linked
with obtaining food. Learning associations between pecking and food come easily to a
pigeon. In contrast, learning to peck to prevent electric shock is not part of the brain
circuitry that a pigeon is born with, and so mastering this association does not come
easily to the bird. Typically, it flies away from noxious situations.
The specific nature of the behavior–consequence associations that animals are able
to learn was first shown in 1966 by psychologist John Garcia. He observed that farmers
in the western United States are constantly shooting at coyotes for attacking lambs;
yet, despite the painful consequences, the coyotes never seem to learn to stop killing
lambs in favor of safer prey. The reason, Garcia speculated, is that a coyote’s brain is
not prewired by adaptation to make this kind of association.
So Garcia proposed an alternative to deter coyotes from killing lambs—one that
uses an association that a coyote’s brain is prepared to make. This association is the
connection between eating something that makes one sick and avoiding that food in
the future. Garcia gave the coyotes a poisoned lamb carcass, which made them sick but
did not kill them.With only one pairing of lamb and illness, most coyotes learned not
to eat sheep for the rest of their lives.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 401
Skinner box
CH11.qxd 2/3/05 4:19 PM Page 401
ls
le
ll
Many humans have similarly acquired food aversions because the taste of a certain
food—especially a novel one—was subsequently paired with illness. This learned taste
aversion is acquired even when the food that was eaten is in fact unrelated to the later
illness. As long as the taste and the nausea are paired in time, the brain is prewired to
make a connection between them.
For instance, one of us ate his first Caesar salad the night before coming down with
a stomach flu. A year later, he was offered another Caesar salad and, to his amazement,
felt ill just at the smell of it. Even though his earlier illness had not been due to the
salad, he had formed an association between the novel flavor and illness.
This kind of strong and rapid associative learning makes adaptive sense. Having a
brain that is prepared to make a connection between a novel taste and subsequent illness
will help an animal avoid poisonous foods and so aid in its survival. Interestingly,
a curious aspect of taste-aversion learning is that we are not even aware of having
formed the association until we encounter the taste again.
The fact that the nervous system is often prewired to make certain associations
but not to make others has led to the concept of preparedness in learning
theories. This concept can help account for some quite complex behaviors. For example,
if two rats are paired in a small box and exposed to a mild electric shock, they
will immediately fight with one another, even though neither was responsible for the
shock.
Apparently, the brain is prepared to make associations between injury and nearby
objects or other animals. Perhaps you have occasionally felt your own temper flare toward
someone who was near you when you were accidentally hurt or in pain for some
reason unrelated to that person. The extent to which we might extend this idea to explain
such human behaviors as bigotry and racism is an interesting topic for debate.
But the point here is that environmental events are working on a brain that is prewired
to make certain types of associations.
Inferring Purpose in Behavior: To Know a Fly
A pitfall in studying the causes of behavior is to infer purpose from an organism’s actions.
In other words, we have a tendency to assume that behavior is intentional. The
problems in making this assumption are illustrated in a wonderful little book titled To
Know a Fly, written by Vincent Dethier.
When a fly lands on a kitchen table, it wanders about, occasionally stomping its
feet. Eventually, it finds a bit of food and sticks its proboscis (a trunklike extension)
into the food and eats. The fly may then walk to a nearby place and begin to groom by
rubbing its legs together quickly. Finally, it spends a long period motionless.
If you observed a fly engaged in these behaviors, it might appear to have been initially
searching for food because it was hungry.When it found food, you might assume
that it gorged itself until it was satisfied, and then it cleaned up and rested. In short, the
fly’s behavior might seem to you to have some purpose or intention.
Dethier studied flies for years to understand what a fly is actually doing when it
engages in these kinds of behaviors. His findings have little to do with purpose or intention.
When a fly wanders about a table, it is not deliberately searching; it is tasting
things that it walks on.
As Figure 11-11 shows, a fly’s taste receptors are on its feet, rather than in its mouth
as in humans. So tasting is automatic when a fly walks. An adult fly’s nervous system
has a built-in preference for sweet tastes and aversions to sour, salty, or bitter flavors.
Therefore, when a fly encounters something sweet, it automatically lowers its proboscis
and eats, or drinks if the sweet is liquid.
402 ! CHAPTER 11
Learned taste aversion. Acquired
association between a specific taste or
odor and illness; leads to an aversion to
foods having that taste or odor.
Preparedness. Predisposition to
respond to certain stimuli differently
from other stimuli.
Crop
Taste receptors
on feet
Midgut
Stretch Foregut
receptors
Brain
Recurrent
nerve
Esophagus
Proboscis
Figure 11-11
Feeding System of the Fly After
having been sampled by taste buds on
the fly’s feet, food is taken in through
the proboscis, after which it passes
through the esophagus to the gut.
Stretch receptors at the entrance to the
gut determine when the esophagus is
full. The recurrent nerve connects with
the brain to signal cessation of eating.
CH11.qxd 2/3/05 4:19 PM Page 402
ls
le
ll
The taste preferences of a fly are interesting. When humans are given very sweet
foods, they normally eat less than they eat of foods that are not as sweet. In contrast,
the sweeter the food is, the more a fly will consume. This taste preference can be measured
by comparing how much a fly drinks of sugar solutions with different concentrations.
The fly has a lower preference for weak sugar solutions and drinks less of them
than very strong solutions.
The fly’s preference for sweet tastes is so great that it will choose food that tastes
very sweet over food that is less sweet but nutritionally better. For instance, when given
a choice between regular glucose and an exceptionally sweet sugar called fucose that a
fly cannot digest, the fly will always choose the fucose, presumably because it tastes better
to a fly. In fact, given the opportunity, a fly will literally die of starvation by eating
nothing but fucose, even though nutritious glucose is available only centimeters away.
Why does a fly stop eating? A logical possibility is that the amount of sugar in its
blood rises to some threshold level. If this possibility were correct, injecting glucose
into the circulatory system of a fly would prevent the fly from eating. But it is not what
happens. Blood-glucose level has no effect on a fly’s feeding. Furthermore, injecting
food into the animal’s stomach or intestine has no effect either. So what is left? The
upper part of the digestive tract.
It turns out that flies have a nerve (the recurrent nerve) that extends from the neck
to the brain and carries information about whether any food is present in the esophagus.
If the recurrent nerve is cut, the fly is chronically “hungry” and never stops eating.
Such flies become so full and fat that their feet no longer reach the ground, and they
become so heavy that they cannot fly.
So what have we learned by studying the fly? The main message is that, even
though a fly appears to act with a “purpose in mind,” a series of very simple mechanisms
actually control its behavior—mechanisms that are not remotely related to our
concept of thought. A fly walks because it is tasting, it eats because its esophagus is devoid
of food, and it stops when its esophagus has some food in it.When the nerve connecting
the esophagus to the brain is cut, a fly will keep on eating even though the food
is flattening its internal organs against the sides of its body.
Hunger is simply the activity of the nerve, not some drive concerning intention.
Clearly, we should not assume that a behavior has a conscious purpose just because it
appears to be the case. Behavior can have very subtle causes that do not include purpose,
which raises the question,How do we know that any behavior is purposeful? That
question turns out to be difficult to answer.
In Review .
Behavior results from evolution and environment, and from the interaction of these two
forces with neurobiological events. Learned behaviors can be selected and influenced by an
individual organism’s own unique experiences as well. The brain of a species is prewired to
produce IRMs to specific sensory stimuli, and prewired brain circuits have been selected by
evolution to prompt associations between certain environmental events. Further, human
taste and smell receptors react chemically with olfactory and gustatory sensations. Their diverse
pathways into the brain eventually merge in the orbitofrontal cortex, leading to the perception
of flavor, the blending of smells and tastes in food. Pheromones are chemosignals
that convey information about the sender and can influence the physiology of the reciever.
In searching for the causes of behavior, then, be aware that behavior can have multiple
causes, which can vary from one behavior to another, and may occur without intent.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 403
CH11.qxd 2/3/05 4:19 PM Page 403
ls
le
ll
404 ! CHAPTER 11
NEUROANATOMY OF MOTIVATED BEHAVIOR
Although the neural circuits that control behavior include regions at all levels of the
brain, the critical structures are the hypothalamus and the associated pituitary gland,
the limbic system, and the frontal lobes. In this section, we investigate the anatomical
and functional organization of these major, functionally interrelated structures.
The hypothalamus receives much of its input from the limbic system and the
frontal lobes, as illustrated in Figure 11-12, where the hypothalamus is represented by
the neck of a funnel. The limbic system and the frontal lobes form the funnel’s rim.
The limbic and frontal regions project to the hypothalamus, which houses many of the
basic circuits and homeostatic mechanisms (processes that keep critical body functions
within a narrow, fixed range) for controlling behavior. To produce behavior, the
hypothalamus sends axons to other brainstem circuits. Thus, although the hypothalamus
plays a central role in controlling motivated behavior, it receives its instructions
from the limbic system and the frontal lobes.
Regulatory and Nonregulatory Behavior
We seek mates, food, or sensory stimulation because of brain activity, but it is convenient
to talk about such behavior as being “motivated.” Like drives, however, motivated
behaviors are not something that we can point to in the brain. Rather,motivations are
inferences that we make about why someone, ourselves included, engages in a particular
behavior. The two general classes of motivated behaviors are regulatory and
nonregulatory. In this section we explore both categories before exploring the neuroanatomy
of motivation.
REGULATORY BEHAVIORS
We explored homeostatic mechanisms regulated by hormones in Chapter 7. Regulatory
behaviors, those motivated by an organism’s survival, are controlled by homeostatic
mechanisms. By analogy, consider a house in which a thermostat is set at 18° Celsius,
like the one in Figure 11-13.When the temperature falls below a certain tolerable range
(say, to 16°C), the thermostat turns the furnace on.When the temperature rises above a
certain tolerable level (say, to 20°C), the thermostat turns on the air conditioner.
Human body temperature is controlled in a somewhat similar manner by a “thermostat”
in the hypothalamus that holds internal temperature at about 37°C, a temperature
Hypothalamus
Behavior
Limbic system Frontal lobes
Figure 11-12
Funneling Signals In this funnel model
of control of motivated behaviors, inputs
from the frontal lobes and limbic system
are funneled through the hypothalamus,
which controls the brainstem circuits to
produce the behavioral patterns.
Some Regulatory Behaviors
Temperature regulation
Eating and drinking
Salt consumption
Waste elimination
Regulatory
system
(thermostat)
When the
temperature falls
below a tolerable
range, the
thermostat causes
the furnace to
turn on.
When the
temperature
rises above a
tolerable range,
the thermostat
causes the air
conditioner to
turn on.
°C
0
10
15 25
5 35
20
30
40
Controlled
systems
Air
conditioner
Furnace
Figure 11-13
Regulatory Mechanism A thermostat
controls temperature inside a house. An
analogous mechanism could control
temperature in the body.
CH11.qxd 2/3/05 4:19 PM Page 404
ls
le
ll
referred to as setpoint. Even slight variations cause us to engage in various behaviors to
regain this setpoint. For example, when body temperature drops slightly, neural circuits
that increase temperature are turned on. These neural circuits might induce an involuntary
response such as shivering or a seemingly voluntary behavior such as moving closer
to a heat source. Conversely, if body temperature rises slightly, we sweat or move to a
cooler place.
Similar mechanisms control many other homeostatic processes, including the
amount of water in the body, the balance of dietary nutrients, and the level of blood
sugar. The control of many of these homeostatic systems is quite complex, requiring
both neural and hormonal mechanisms.However, in some way, all of the body’s homeostatic
systems include the activity of the hypothalamus.
Imagine that specific cells are especially sensitive to temperature. When they are
cool, they become very active; when they are warm, they become less active. These cells
could function as a thermostat, telling the body when it is too cool or too warm.A similar
set of cells could serve as a “glucostat,” controlling the level of sugar in the blood,
or as a “waterstat,” controlling the amount of H2O in the body. In fact, the body’s real
homeostatic mechanisms are slightly more complex than this imagined one, but they
work in accord with the same general principle.
We evolved mechanisms to hold conditions such as temperature constant because
the body, including the brain, is a chemical “soup” in which thousands of chemical reactions
are taking place all the time.Maintaining constant temperature becomes critical.
When temperature changes, even by such a small amount as 2 Celsius degrees, the
rates at which chemical reactions take place change.
Such changes might be tolerable, within certain limits, if all the reaction times
changed to the same extent. But they do not do so. Consequently, an increase of 2 degrees
might increase one reaction by 10 percent and another by only 2 percent. Such
uneven changes would wreak havoc with finely tuned body processes such as metabolism
and the workings of neurons.
A similar logic applies to maintaining homeostasis in other body systems. For
instance, cells require certain concentrations of water, salt, or glucose to function properly.
If those concentrations were to fluctuate wildly, they would cause a gross disturbance
of metabolic balance and a subsequent biological disaster.
NONREGULATORY BEHAVIORS
In contrast with regulatory behaviors, such as eating or drinking, nonregulatory
behaviors are neither required to meet the basic needs of an animal nor controlled by
homeostatic mechanisms. Thus, nonregulatory behaviors include everything else that
we do—from sexual intercourse to parenting to such curiosity-driven activities as conducting
psychology experiments. Some nonregulatory behaviors, such as sexual intercourse,
entail the hypothalamus, but most of them probably do not. Rather, such
behaviors entail a variety of forebrain structures, especially the frontal lobes. Presumably,
as the forebrain evolved to a larger size, so did our range of nonregulatory
behaviors.
Most nonregulatory behaviors are strongly influenced by external stimuli. As a result,
sensory systems must play some role in controlling them. For example, the sexual
behavior of most male mammals is strongly influenced by the pheromone emitted by
receptive females. If the olfactory system is not functioning properly, we can expect
abnormalities in sexual behavior. We will return to the topic of sexual behavior later
in this chapter when we investigate it as an example of how a nonregulatory behavior
is controlled. But first we will look at the brain structures taking part in motivated
behaviors—both nonregulatory and regulatory ones.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 405
Homeostatic mechanism. Process that
maintains critical body functions within a
narrow, fixed range.
Regulatory behavior. Behavior
required to meet the basic needs of the
animal.
Nonregulatory behavior. Behavior not
required to meet the basic needs of the
animal.
Some Nonregulatory Behaviors
Sexual behavior
Parental behavior
Aggression
Food preference
Curiosity
Reading
CH11.qxd 2/3/05 4:19 PM Page 405
ls
le
406 ! CHAPTER 11
The Hypothalamic Circuit’s Relaying Function
One function of the hypothalamus, as stated earlier, is to regulate our internal environment.
It maintains homeostasis by acting on both the endocrine system and the autonomic
nervous system (ANS). The hypothalamus also influences the selection of
behaviors by the rest of the brain, especially by the limbic system, as you will discover
later in the chapter when we consider emotional behavior. In these ways, the hypothalamus,
although it constitutes less than 1 percent of the human brain’s volume, controls
an amazing variety of behaviors, ranging from heart rate to feeding to sexual
activity.
HYPOTHALAMIC INVOLVEMENT IN
HORMONE SECRETIONS
A principal function of the hypothalamus is to control the pituitary gland, which is
attached to the hypothalamus by a stalk (Figure 11-14A). The optic nerves cross to
form the optic chiasm right in front of the hypothalamus, and the optic tracts are just
lateral to it (Figure 11-14B).
The hypothalamus can be divided into three regions: the lateral, the medial, and the
periventricular, illustrated in frontal section in Figure 11-14B. The lateral hypothalamus
is composed both of nuclei and of tracts running up and down the brain, connecting
the lower brainstem to the forebrain. The principal tract, shown in Figure 11-15, is the
medial forebrain bundle (MFB). The MFB, which connects structures in the brainstem
Figure 11-14
The Nuclei and Regions of the
Hypothalamus (A) Medial view shows
the relation between the hypothalamic
nuclei and the rest of the brain. (B) In
frontal section, the relation between
the hypothalamus and the third
ventricle can be seen. The three
principal hypothalamic regions are the
periventricular, lateral, and medial.
On the CD, examine the
hypothalamus in three dimensions in the
subsection on subcortical structures in the
module on the Central Nervous System.
Pituitary gland. Endocrine gland
attached to the bottom of the
hypothalamus; its secretions control
the activities of many other endocrine
glands; known to be associated with
biological rhythms.
Medial forebrain bundle (MFB).
Tract that connects structures in the
brainstem with various parts of the limbic
system; forms the activating projections
that run from the brainstem to the basal
ganglia and frontal cortex.
Ventromedial
hypothalamic
nucleus
Posterior
nucleus
Preoptic
nucleus
Paraventricular
nucleus
Dorsomedial
hypothalamic nucleus
Pituitary
stalk
Pituitary
gland
(A)
(B)
Third ventricle
Medial
hypothalamic
region
Optic tract
Ventromedial
hypothalamic
nucleus
Periventricular
region
Lateral
hypothalamic
region
Hypothalamus
CH11.qxd 2/3/05 4:19 PM Page 406
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 407
with various parts of the limbic system, forms the activating projections that run
from the brainstem to the basal ganglia and frontal cortex.
Fibers that ascend from the dopamine- and noradrenaline-containing cells of
the lower brainstem form a significant part of the MFB. Recall from Chapter 7
that dopamine is important in our experience of reward. Thus, the dopaminecontaining
fibers of the MFB contribute to the control of many motivated behaviors,
including eating and sexual behaviors.
Each nucleus of the hypothalamus is anatomically distinct, but most have
multiple functions. These multiple functions are due, in part, to the fact that the
cells in different nuclei contain various peptide transmitters, each of which plays a
role in different behaviors. For instance, the transmitters in the cells in the paraventricular
nucleus may be vasopressin, oxytocin, or various combinations of
other peptides (such as enkephalin and neurotensin).
The production of various neuropeptides hints at the special relation between the
hypothalamus and the pituitary. The pituitary consists of distinct anterior and posterior
glands, as shown in Figure 11-16. The posterior pituitary is composed of neural
tissue and is essentially a continuation of the hypothalamus.
Neurons in the hypothalamus make peptides (e.g., oxytocin and vasopressin)
that are transported down their axons to terminals lying in the posterior pituitary. If
these neurons become active, they send action potentials to the terminals, causing the
Frontal
cortex Hypothalamus
Medial
forebrain bundle
Temporal
lobe
Basal ganglia Limbic cortex
Figure 11-15
Medial Forebrain Bundle Major
components of the MFB, a major
pathway for fibers connecting various
parts of the limbic system with the
brainstem, are the activating projections
that run from the brainstem to the basal
ganglia and frontal cortex.
Figure 11-16
Hypothalamus and Pituitary
Gland The pituitary has two
divisions: the anterior and
posterior. The anterior
pituitary is connected to the
hypothalamus by a system of
blood vessels that carry
hormones from the
hypothalamus to the pituitary.
The posterior pituitary receives
input from axons of
hypothalamic neurons. Both
regions of the pituitary respond
to hypothalamic input by
producing hormones that travel
in the bloodstream to stimulate
target organs.
Blood in
Blood in
Capillaries
Axon
terminals
Posterior
pituitary
Hypothalamus
Anterior
pituitary
Blood out Blood out
(carrying posterior
pituitary hormones)
4
Releasing hormones then
leave the capillaries and act
on hormone-secreting
anterior pituitary cells.
2
Hormones released by axon
terminals in the posterior
pituitary are picked up by
capillaries and carried into
the bloodstream.
Pituitary
gland
1
Hormones are
synthesized here and
sent to axon terminals
in the posterior
pituitary.
3
Releasing hormones are
synthesized here and
secreted into capillaries that
carry them to the anterior
pituitary.
CH11.qxd 2/3/05 4:19 PM Page 407
Major Hormones Produced by the Anterior Pituitary
Hormone Function
Adrenocorticotrophic hormone (ACTH) Controls secretions of the adrenal cortex
Thyroid-stimulating hormone (TSH) Controls secretions of the thyroid gland
Follicle-stimulating hormone (FSH) Controls secretions of the gonads
Luteinizing hormone (LH) Controls secretions of the gonads
Prolactin Controls secretions of the mammary glands
Growth hormone (GH) Promotes growth throughout the body
Table 11-1
ls
le
ll
terminals to release the peptides that are stored there. But rather than affecting another
neuron, as occurs at most synapses, these peptides are picked up by capillaries in the
posterior pituitary’s rich vascular bed.
From there, they enter the body’s bloodstream. The blood then carries the peptides
to distant targets, where they have their effects. For example, vasopressin affects water
resorption by the kidneys, and oxytocin controls both uterine contractions and the
ejection of milk by mammary glands in the breasts. Peptides can have multiple functions,
depending on the location of receptors. Thus, oxytocin not only controls milk
ejection in females but also plays a more general role in several forms of affiliative behavior,
including parental care, grooming, and sexual behavior in both men and
women (Insel & Fernald, 2004).
The glandular tissue of the anterior pituitary synthesizes various hormones, the
major ones being listed in Table 11-1.The hypothalamus controls the release of these anterior
pituitary hormones by producing chemicals known as releasing hormones. Produced
by hypothalamic cell bodies, releasing hormones are secreted into tiny blood
vessels,or capillaries, that transport them to the anterior pituitary, as Figure 11-16 shows.
A releasing hormone can either stimulate or inhibit the release of an anterior pituitary
hormone. For example, the hormone prolactin is produced by the anterior
pituitary, but its release is controlled by a prolactin-releasing factor and a prolactin release–
inhibiting factor, both synthesized in the hypothalamus. The release of hormones
by the anterior pituitary in turn provides a means by which the brain can control what
is taking place in many other parts of the body. Three factors control hypothalamic
hormone-related activity: feedback loops, neural regulation, and responses based on
experience.
Feedback Loops When the level of, say, thyroid hormone is low, the hypothalamus releases
thyroid-stimulating hormone-releasing hormone (TSH-releasing hormone). The
TSH-releasing hormone stimulates the anterior pituitary to release thyroid-stimulating
hormone, which then acts on the thyroid gland to secrete more thyroid hormone.
There must, however, be some control over how much thyroid hormone is secreted,
and so the hypothalamus has receptors to detect the level of thyroid hormone.
When that level rises, the hypothalamus lessens its secretion of TSH-releasing hormone.
This type of system is essentially a form of homeostatic control that works as a
feedback mechanism, a system in which a neural or hormonal loop regulates the activity
of neurons, initiating the neural activity or hormone release, as illustrated in Figure
11-17A.
The hypothalamus initiates a cascade of events that result in the secretion of hormones,
but it pays attention to how much hormone is released.When a certain level is
reached, it stops its hormone-stimulating signals. Thus, the feedback mechanism in the
hypothalamus maintains a fairly constant circulating level of certain hormones.
408 ! CHAPTER 11
Releasing hormones. Peptides that are
released by the hypothalamus and act to
increase or decrease the release of
hormones from the anterior pituitary.
Thyroid
gland
CH11.qxd 2/3/05 4:19 PM Page 408
ls
le
ll
Neural Control A second control over
hormone-related activities of the hypothalamus
requires regulation by other brain structures,
such as the limbic system and the frontal lobes.
Figure 11-17B diagrams this type of control in
relation to the effects of oxytocin released from
the paraventricular nucleus of the hypothalamus.
As stated earlier, one function of oxytocin is to
stimulate cells of the mammary glands to release
milk. As an infant suckles the breast, the tactile
stimulation causes hypothalamic cells to release
oxytocin, which stimulates milk letdown. In this
way, the oxytocin cells participate in a fairly simple
reflex that is both neural and hormonal.
Other stimuli also can influence the release
of oxytocin, however, which is where control by
other brain structures comes in. For example, the sight, sound, or even thought of her
baby can trigger a lactating mother to eject milk. Conversely, as diagrammed in Figure
11-17B, feelings of anxiety in a lactating woman can inhibit milk ejection. These excitatory
and inhibitory influences exerted by cognitive activity imply that the cortex can
influence neurons in the paraventricular region. Projections from the frontal lobes to
the hypothalamus likely perform this role.
Experiential Responses A third way in which the hormone-related activities of the
hypothalamus are controlled is by the brain’s responses to experience. In response to
experience, neurons in the hypothalamus undergo structural and biochemical changes
just as cells in other brain regions do. In other words, hypothalamic neurons are like
neurons elsewhere in the brain in that they can be changed by prolonged demands
placed on them.
Such changes in hypothalamic neurons can affect the output of hormones. For instance,
when a woman is lactating, the cells producing oxytocin increase in size to promote
oxytocin release to meet the increasing demands of a growing infant for more
milk. Through this control, which is mediated by experience, the baby is provided with
sufficient milk over time.
HYPOTHALAMIC INVOLVEMENT IN
GENERATING BEHAVIOR
So far, we have considered the role of the hypothalamus in controlling hormone systems,
but equally important is its role in generating behavior. This function was first
demonstrated by studies in which stimulating electrodes were placed into the hypothalami
of various animals, ranging from chickens to rats and cats.When a small electric
current was delivered through a wire electrode, an animal suddenly engaged in
some complex behavior. The behaviors included eating and drinking; digging; and displaying
fear, attack, predatory, or reproductive behavior.
The particular behavior depended on which of many sites in the hypothalamus
was stimulated. All the behaviors were smooth, well integrated, and indistinguishable
from normally occurring ones. Furthermore, all were goal directed.
The onset and termination of these behaviors depended entirely on the hypothalamic
stimulation. For example, if an electrode in a certain location elicited feeding behavior,
the animal would eat as soon as the stimulation was turned on and would
continue to eat until the stimulation was turned off. If the food was removed, however,
the animal would neither eat nor engage in other behaviors such as drinking. Recall
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 409
(A) Feedback loops
Hypothalamus Hypothalamus
Releasing
hormones
(+)
Feedback
Hormones
Anterior
pituitary
Target
organs
Anxiety
Mammary glands
Milk letdown
Oxytocin
(B) Milk-letdown response
(–) Infant-related (+) (–)
stimuli
Pituitary
hormones
(+)
Figure 11-17
Hypothalamic Controls (A) The
hormone-controlling feedback loops
from the hypothalamus to the pituitary
and its target organs. The hypothalamus
releases hormones, which stimulate
the anterior pituitary to release its
hormones, which stimulate target
organs such as the thyroid or adrenal
gland to release their hormones. Those
hormones act, in turn, to influence the
hypothalamus to decrease its secretion
of the releasing hormone. (B) Oxytocin
stimulates the mammary glands to
release milk. Oxytocin release from the
hypothalamus is enhanced by infantrelated
stimuli and inhibited by maternal
anxiety.
CH11.qxd 2/3/05 4:19 PM Page 409
ls
le
ll
410 ! CHAPTER 11
that Roger ate continuously if foodlike materials were present, corresponding to the
continuous activity of his hypothalamus caused by the tumor.
Figure 11-18 illustrates the effect of stimulation at a site that elicits digging.When
there is no current, the animal sits quietly.When the current is turned on, the animal
digs into the sawdust vigorously; when the current is turned off, the animal stops digging.
If the sawdust is removed, there also is no digging.
Two important additional characteristics of the behaviors generated by hypothalamic
stimulation are related to (1) the survival of the animal and the species and (2)
reward. Animals apparently find the stimulation of these behaviors pleasant, as suggested
by the fact that they willingly expend effort, such as pressing a bar, to trigger the
stimulation. Recall that cats kill birds and mice because the act of stalking and killing
prey is rewarding to them. Similarly, we can hypothesize that animals eat because
eating is rewarding, drink because drinking is rewarding, and mate because mating is
rewarding.
The Limbic Circuit’s Organizing Function
We now turn our attention to parts of the brain that interact with the hypothalamus
in generating motivated behaviors. These brain structures evolved as a ring of structures
around the brainstem in early amphibians and reptiles.Nearly 150 years ago, Paul
Broca was impressed by this evolutionary development and called these structures the
“limbic lobe” (from the Latin word limbus, meaning “border” or “hem”).
Known collectively as the limbic system today, these structures are actually a primitive
cortex, as described in Chapter 2. In mammals, the limbic cortex encompasses the
cingulate gyrus and the hippocampal formation, as shown in Figure 11-19. The hippocampal
formation includes the hippocampus, a cortical structure that plays a role
in species-specific behaviors, memory, and spatial navigation and is vulnerable to the
effects of stress, and the parahippocampal cortex adjacent to it.
ORGANIZATION OF THE LIMBIC CIRCUIT
As anatomists began to study the limbic-lobe structures, connections between these
structures and the hypothalamus became evident. It also became apparent that the
limbic lobe has a role in emotion. For instance, in the 1930s, James Papez observed
that people with rabies had radical abnormalities in their emotional behavior, and
In the absence of
stimulation, the animal
sits quietly.
When stimulated, the
animal digs vigorously.
The animal stops digging
when stimulation stops.
Stimulation
wire
Figure 11-18
Generating Behavior When rats
receive electrical stimulation to the
hypothalamus, they produce goaldirected
behaviors. This rat is stimulated
to dig when and only when the
electricity is turned on. Note also that,
if the sawdust is removed (not shown in
the bottom drawing), there is no
digging.
Temporal
lobe
Corpus callosum
Fornix nucleus of
dorsal
thalamus
Anterior
Hippocampal
formation
Mammillothalamic
tract
Olfactory
bulb
Basal
forebrain
nuclei
Amygdala
Mammillary
body
Hippocampus
Parahippocampal
cortex
Cingulate gyrus
cortex
Prefrontal
Figure 11-19
Limbic Lobe Encircling the brainstem, the limbic lobe as described by Broca
consists of the cingulate gyrus and hippocampal formation (the hippocampus
and parahippocampal cortex), the amygdala, the mammillothalamic tract, and
the anterior thalamus.
CH11.qxd 2/3/05 4:20 PM Page 410
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 411
postmortems showed that the rabies had selectively
attacked the hippocampus. (The definitive proof
of rabies is still a postmortem examination of the
hippocampus.)
On the basis of this finding, Papez concluded that
the limbic lobe and associated subcortical structures
provide the neural basis of emotion. He proposed a
circuit, traced in Figure 11-20A, now known as the
Papez circuit, whereby emotion could reach consciousness, which was presumed to reside
in the cerebral cortex. Papez’s limbic-circuit concept (also called the limbic system)
was expanded by Paul MacLean in 1949 to include the amygdala and prefrontal
cortex as well. Figures 11-19 and 11-20A show the amygdala lying adjacent to the hippocampus
in the temporal lobe, with the prefrontal cortex lying just anterior.
Figure 11-20B charts the limbic circuit schematically. The hippocampus, amygdala,
and prefrontal cortex all connect with the hypothalamus. The mammillary nucleus
of the hypothalamus connects to the anterior thalamus, which in turn connects
with the cingulate cortex, which then completes the circuit by connecting with the hippocampal
formation, amygdala, and prefrontal cortex. This anatomical arrangement
can be compared to the funnel in Figure 11-20C, which shows the hypothalamus as the
spout leading to motivated behavior.
There is now little doubt that most structures of the limbic system, especially the
amygdala and hypothalamus, take part in emotional behaviors, as detailed later in the
chapter. But most limbic structures are now known to play an important role in various
motivated behaviors as well as in emotion, especially in motivating species-typical
behaviors such as feeding and sexual activity. The critical structures for such motivated
behaviors, as well as for emotion, are the amygdala and the hypothalamus. Having already
considered the hypothalamus, we now turn to the amygdala.
THE AMYGDALA
Named for the Greek word for “almond,” the amygdala consists of three principal subdivisions:
the corticomedial area, the basolateral area, and the central area. Like the hypothalamus,
the amygdala receives inputs from all sensory systems. But, in contrast
with the neurons of the hypothalamus, those of the amygdala require more-complex
stimuli to be excited.
In addition, many amygdala neurons are multimodal: they respond to more than
one sensory modality. In fact, some neurons in the amygdala respond to sight, sound,
touch, taste, and smell stimuli. These cells must create a rather complex image of the
sensory world.
Hippocampus. Distinctive, threelayered
subcortical structure of the limbic
system lying in the medial region of the
temporal lobe; plays a role in speciesspecific
behaviors, memory, and spatial
navigation and is vulnerable to the effects
of stress.
Amygdala. Almond-shaped collection
of nuclei located within the limbic
system; plays a role in emotional and
species-typical behaviors.
On the CD, see the hippocampus and
the amygdala in three dimensions in the
subsection on subcortical structures in the
module on the Central Nervous System.
Prefrontal
cortex
Prefrontal
cortex
Sensory association
cortex
Anterior
thalamus
Anterior
thalamus
Hypothalamus
Hypothalamus
Sensory
association
cortex
Hippocampal
formation
Hippocampal
formation
Amygdala
Amygdala
Cingulate
cortex
Cingulate
cortex
(A) (B)
Behavior
(C)
Hypothalamus
Behavior
lobes
system
Frontal
Limbic
Mammillary
nucleus
Mammillary
nucleus
Figure 11-20
Limbic System (A) In this
modern conception of the limbic
system, an interconnected
network of structures, the Papez
circuit, controls emotional
expression. (B) A schematic
representation, coded to brain
areas shown in part A by color,
charting the major connections
of the limbic system. (C) A
reminder that parts A and B can
be conceptualized as a funnel of
outputs through the
hypothalamus.
CH11.qxd 2/3/05 4:20 PM Page 411
ls
le
ll
The amygdala sends connections primarily to the hypothalamus and the brainstem,
where it influences neural activity associated with emotions and species-typical
behavior. For example, when the amygdalae of epileptic patients are electrically stimulated
before brain surgery, the patients become fearful and anxious. We observed a
woman who responded with increased respiration and heart rate, saying that she felt
as if something bad was going to happen, although she could not specify what.
Amygdala stimulation can also induce eating and drinking. We observed a man
who drank water every time the stimulation was turned on. (There happened to be a
pitcher of water on the table next to him.) Within 20 minutes, he had consumed about
2 liters of water. When asked if he was thirsty, he said, “No, not really. I just feel like
drinking.”
The amygdala’s role in eating can be seen in patients with lesions in the amygdala.
These patients, much as we saw in the effects of Roger’s tumor, may be much less discriminating
in their food choices, eating foods that were formerly unpalatable to them.
Lesions of the amygdala may also give rise to hypersexuality.
The Frontal Lobes’ Executive Function
The amygdala is intimately connected with the functioning of the frontal lobes that
constitute all cortical tissue in front of the central sulcus. This large area is made up of
several functionally distinct cortical regions. Figure 11-21 shows the three main regions:
the motor cortex, the premotor cortex, and the prefrontal cortex.
As you learned in Chapter 10, the motor cortex controls fine movements,
especially of the fingers, hands, toes, feet, tongue, and face. The premotor
cortex participates in the selection of appropriate movement
sequences. For instance, a resting dog may get up in response to its owner’s
call, which serves as an environmental cue for a series of movements
processed by one region of the premotor cortex. Or a dog may get up for
no apparent reason and wander about the yard, which is a sequence of actions
in response to an internal cue, this time processed by a different region
of the premotor cortex.
Finally, the prefrontal cortex (which literally means “in front of the
front”) is anterior to the premotor cortex. It is made up of two primary
areas: the dorsolateral region and the inferior region. Recall from Chapter 10 that the
prefrontal cortex plays a role in specifying the goals toward which movement should
be directed. In this role, it controls the processes by which we select movements that
are appropriate for the particular time and place. This selection may be cued by internal
information (such as memory and emotion) or it may be made in response to context
(environmental information).
Like the amygdala, the frontal lobes, particularly the prefrontal cortex, receive
highly processed information from all sensory areas. Many of the neurons in the prefrontal
cortex, like those in the amygdala, are multimodal. As shown in Figure 11-22,
the prefrontal cortex receives connections from the amygdala, the dorsomedial thalamus,
the posterior parietal (sensory association) cortex, and the dopaminergic cells of
the ventral tegmental area.
The dopaminergic input plays an important role in regulating how prefrontal neurons
react to stimuli, including emotional ones. Abnormalities in this dopaminergic
projection may account for some disorders, including schizophrenia, in which people
have little emotional reaction to normally arousing stimuli (see Chapter 15).
Figure 11-22 also shows the areas to which the prefrontal cortex sends connections.
The inferior prefrontal region projects axons to the amygdala and the hypothalamus in
412 ! CHAPTER 11
Prefrontal cortex. The cortex lying in
front of the motor and premotor cortex of
the frontal lobe; the prefrontal cortex is
particularly large in the human brain.
Inferior
prefrontal
cortex
Dorsolateral
prefrontal
cortex
Premotor
cortex
Orbital
cortex
Motor
cortex
Central
sulcus
Prefrontal
cortex
Figure 11-21
Lateral View of the Gross Subdivisions
of the Frontal Lobe
CH11.qxd 2/3/05 4:20 PM Page 412
ls
le
ll
particular. These axons provide a route for influencing the autonomic
system, which controls changes in blood pressure, respiration,
and so forth. The dorsolateral prefrontal region sends its
connections primarily to the posterior parietal cortex, the cingulate
cortex, the basal ganglia, and the premotor cortex. These connections
provide a route for influencing movement as well as certain
memory functions to be considered in Chapter 13.
As already stated, the prefrontal cortex takes part in the selection
of behaviors appropriate to the particular time and place. This
selection may be cued by internal information or it may be made in
response to the environmental context. Disruption to this selection
function can be seen in people with injury to the dorsolateral frontal
lobe. They become overly dependent on environmental cues to determine
their behavior. Like small children, they can be easily distracted
by what they see or hear.
We have all experienced this kind of loss of concentration to
some extent, but, for a frontal-lobe patient, the problem is exaggerated
and persistent. Because the person becomes so absorbed in irrelevant
stimuli, he or she is unable to act on internalized information
most of the time.
A good example is J. C., whose bilateral damage to the dorsolateral
prefrontal cortex resulted from having a tumor removed. J. C.
would lie in bed most of the day fixated on television programs. He
was aware of his wife’s opinion of this behavior, but only the opening
of the garage door when she returned home from work in the evening
would stimulate him into action.Getting out of bed was controlled by this specific environmental
cue and, without it, he seemed to lack motivation. Television completely distracted
him from acting on internal knowledge of things that he could or should do.
Adapting behavior appropriately to the environmental context also is a function of
the prefrontal cortex.Most people readily change their behavior to match the situation
at hand. We behave in one way with our parents, in another with our friends, in another
with our children, and in yet another with our coworkers. Each set of people creates
a different context, and we shift our behaviors accordingly. Our tone of voice, our
use of slang or profanity, and the content of our conversations are quite different in
different contexts.
Even among our peers, we act differently, depending on who is present.We may be
relaxed in the presence of some people and ill at ease with others. It is therefore no accident
that the size of the frontal lobes is related to the sociability of a species’ behavior.
Social behavior is extremely rich in contextual information, and humans are highly
social.
The control of behavior in context requires detailed sensory information, which is
conveyed from all the sensory regions to the frontal lobes. This sensory input includes
not only information from the external world but internal information from the ANS
as well. People with damage to the inferior prefrontal cortex, which is common in traumatic
brain injuries, have difficulty adapting their behavior according to the context,
especially the social context. Consequently, they often make social gaffes.
In summary, the role of the frontal lobes in selecting behaviors is important in considering
behavioral causes.The frontal lobes act much like a composer but, instead of selecting
notes and instruments, they select our actions.Not surprisingly, the frontal lobes
are sometimes described as housing executive functions, a concept that we will return to
in Chapter 14 in considering the frontal lobe’s role in planning.To grasp the full extent of
frontal-lobe control of behavior, see “Agenesis of the Frontal Lobe” on page 414.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 413
Dorsomedial
thalamus
Inputs to
prefrontal cortex
Outputs from
prefrontal cortex
Amygdala
Sensory association
cortex
Ventral tegmental
area (dopamine)
Hypothalamus Cingulate
cortex
Posterior
parietal
cortex
Premotor
cortex
Basal
ganglia
Prefrontal
cortex
Click on the area on the Central
Nervous System on your CD to see a
model and sections of the frontal lobes.
Figure 11-22
Prefrontal Connections The prefrontal
cortex receives inputs from all of the
sensory systems, the amygdala, the
dorsal medial thalamus, and the
dopamine-rich cells of the ventral
tegmentum. The prefrontal cortex
sends connections to the amygdala,
premotor cortex, basal ganglia, posterior
parietal cortex, and hypothalamus.
CH11.qxd 2/3/05 4:29 PM Page 413
ls
le
ll
In Review .
Motivated behaviors appear to be goal directed and purposeful. Regulatory behavior is controlled
by a homeostatic mechanism that works to keep a vital aspect of body function
within a narrow, fixed range. Nonregulatory behaviors consist of everything else that we
do. Many nonregulatory behaviors are partly controlled by external stimuli that serve as
cues. Within the brain, the hypothalamus, the limbic system, and the frontal lobes house
the major behavioral circuitry. The hypothalamus provides the simplest, largely homeostatic,
control. The limbic system stimulates emotional reactions and species-typical behaviors,
whereas the frontal lobes generate the rationale for behavior at the right time and
place, taking factors such as external events and internal information into account.
Agenesis of the Frontal Lobes
Focus on Disorders
The role of the frontal lobes in motivated behavior is perhaps
best understood by looking at J. P.’s case, described in detail
by Stafford Ackerly (1964). J. P., who was born in December
1914, was a problem child. Early on, he developed the habit
of wandering. Policemen would find him miles from home,
as he had no fear of being lost. Severe whippings by his father
did not deter him.
J. P.’s behavioral problems continued as he grew older,
and, by adolescence, he was constantly in trouble. Yet J. P.
also had a good side. When he started school, his first-grade
teacher was so impressed with his polite manners that she
began writing a letter to his parents to compliment them on
having such a well-mannered child who was such a good influence
in the class.
As she composed the letter, she looked up to find J. P.
exposing himself to the class and masturbating. This contradiction
of polite manners and odd behavior characterized J.
P.’s conduct throughout his life. At one moment, he was
charming and, at the next, he was engaged in socially unacceptable
behavior.
He developed no close friendships with people of either
sex, in large part because of his repeated incidents of public
masturbation, stealing, excessive boastfulness, and wandering.
He was a person of normal intelligence who seemed unaffected
by the consequences of his behavior. Police officers,
teachers, and neighbors all felt that he was willfully behaving
in an asocial manner and blamed his parents for not enforcing
strict enough discipline.
Perhaps as a result, not until he was 19 years old was
J. P.’s true condition detected. To prevent J. P. from serving a
prison term for repeated automobile theft, a lawyer suggested
that J. P. undergo psychiatric evaluation. He was examined by
a psychiatrist, who ordered a brain scan. The image revealed
that J. P. lacked a right frontal lobe. Furthermore, his left
frontal lobe was about 50 percent of normal size.
It is almost certain that he simply never developed
frontal lobes. The failure of a structure to develop is known
as agenesis; J. P. had agenesis of the frontal lobes. His case
offers an unusual opportunity to study the role of the frontal
lobes in motivated behavior.
Clearly, J. P. lacked the “bag of mental tricks” that most
people use to come to terms with the world. Normally, behavior
is affected both by its past consequences and by current
environmental input. J. P. did not seem much influenced
by either of these factors. As a result, the world was simply
too much for him.
He always acted childlike and was unable to formulate
plans for the future or to inhibit many of his behaviors. He
acted on impulse. At home, he was prone to aggressive outbursts
about small matters, especially with regard to his
mother.
Curiously, he seemed completely unaware of his life situation.
Even though the rest of his brain was working fairly
well (his IQ was normal and his language skills were very
good), the functional parts of his brain were unable to compensate
for the absence of the frontal lobes.
CH11.qxd 2/3/05 4:20 PM Page 414
ls
le
ll
STIMULATING EMOTION
Even though we all know what emotions are, it is easier to identify how emotions are
expressed than it is to define them. The concept of emotion is difficult to define because
emotion, like motivation, is intangible; it is an inferred state. The expression of
emotions includes physiological changes, in heart rate, blood pressure, and hormone
secretions. It also includes certain motor responses, especially the movements of the
facial muscles that produce facial expression (see Figure 11-5).
The importance of emotion to our everyday lives cannot be underestimated.
Emotion, for example, is the inspiration for artistic expression ranging from poetry
to filmmaking to painting. Indeed, one reason that many people enjoy the arts is that
they evoke emotions. We can therefore conclude that people find certain emotions
pleasant. On the other hand, severe and prolonged negative emotions, especially
anxiety and depression, can cause clinical disorders. Because so much of human
life revolves around emotions, understanding them is central to understanding our
humanness.
To explore the neural control of emotions, we must first specify the types of behavior
that we want to explain. Think of any significant emotional experience that you
had recently. Perhaps you had a serious disagreement with a close friend. Maybe you
just became engaged to be married.
A common characteristic of such experiences is that they include autonomic responses
such as rapid breathing, sweating, and dry mouth. They may also entail strong
subjective feelings that we often label as anger, fear, or love. Finally, emotions typically
entail thoughts or plans related to the experience, which may take the form of replaying
conversations and events in your mind, anticipating what you might say or do
under similar circumstances in the future, or planning your married life.
These three forms of experience suggest the influence of different neural systems.
The autonomic component must include the hypothalamus and associated
structures. The feelings are more difficult to localize but clearly include the amygdala
and probably parts of the frontal lobes. And, finally, the cognitions are likely to
be cortical.
What is the relation between our cognitive experience of an emotion and the
physiological changes associated with it? One view is that the physiological changes
(such as trembling and rapid heartbeat) come first, and the brain then interprets these
changes as an emotion of some kind. This perspective implies that the brain (most
likely the cortex) creates a cognitive response to autonomic information.
That response varies with the context in which the autonomic arousal occurs. For
example, if we are frightened by a movie, we experience a weaker, more short-lived
emotion than if we are frightened by a real-life encounter with a gang of muggers.Variations
of this perspective have gone by many names, beginning with the James-Lange
theory, named for its originators. All assume that the brain concocts a story to explain
bodily reactions.
Two lines of evidence support the James-Lange theory and similar points of view.
One is that the same autonomic responses can accompany different emotions. In other
words, particular emotions are not tied to their own unique autonomic changes. This
line of evidence leaves room for interpreting what a particular pattern of arousal
means, even though particular physiological changes may suggest only a limited range
of possibilities. The physiological changes experienced during fear and happiness are
unlikely to be confused with one another, for instance.
The second line of evidence supporting the view that physiological changes are the
starting point for emotions comes from people with reduced information about their
own autonomic arousal, owing to spinal-cord injury, for example. Such people suffer
a decrease in perceived emotion, the severity of which depends on how much sensory
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 415
CH11.qxd 2/3/05 4:20 PM Page 415
ls
le
ll
input they have lost. Figure 11-23 illustrates this relation. It shows that people
with the greatest loss of sensory input, which occurs with injuries at the uppermost
end of the spinal cord, also have the greatest loss of emotional intensity.
In contrast, people with low spinal injuries retain most of their visceral
input and have essentially normal emotional reactions.
Antonio Damasio (1999) emphasized an important additional aspect
of the link between emotion and cognitive factors in his somatic marker
hypothesis.When Damasio studied patients with frontal-lobe injuries, he was
struck by how they could be highly rational in analyzing the world yet still
make decidedly irrational social and personal decisions. The explanation, he
argued, is that the reasoning of people with frontal-lobe injury is no longer affected,
either consciously or unconsciously, by the neural machinery that underlies
emotion. Cut off from critical emotional input, many social and
personal decisions are therefore rather poor.
To account for these observations, Damasio proposed that emotions are
responses induced by either internal or external stimuli not normally attended
to consciously. For example, if you encounter a bear as you walk down the
street, presuming that you live in a place where this event could take place, the
stimulus is processed rapidly without conscious appraisal. In other words, a
sensory representation of the bear in the visual cortex is transmitted directly
to brain structures, such as the amygdala, that initiate an emotional response.
This emotional response includes actions on structures in the forebrain
and brainstem and ultimately on the autonomic nervous system. As mentioned
earlier, the amygdala has connections to the frontal lobes, and so the
emotional response can influence the frontal lobes’ appraisal of the world.
However, if the frontal lobes are injured, the emotional information is excluded
from cognitive processing, and so the quality of emotion-related appraisals
suffers. In other words, the response to the bear might be inappropriate.
To summarize, Damasio’s somatic marker hypothesis proposes how emotions are
normally linked to a person’s thoughts, decisions, and actions. In a typical emotional
state, certain regions of the brain send messages to many other brain areas and to most
of the rest of the body through hormones and the ANS. These messages produce a
global change in the organism’s state, and this altered state influences behavior, usually
in a nonconscious way.
The Amygdala and Emotional Behavior
In addition to controlling certain species-typical behaviors, the amygdala influences
emotion (Davis et al., 2003). The role of the amygdala can be seen most clearly in monkeys
whose amygdalae have been removed. In 1939, Heinrich Klüver and Paul Bucy
reported an extraordinary result, now known as the Klüver-Bucy syndrome, that followed
the removal of the amygdalae and anterior temporal cortices of monkeys. The
principal symptoms include:
1. tameness and loss of fear;
2. indiscriminate dietary behavior (eating many types of formerly rejected foods);
3. greatly increased autoerotic, homosexual, and heterosexual activity, with inappropriate
object choice (e.g., the sexual mounting of chairs);
4. tendency to attend to and react to every visual stimulus;
5. tendency to examine all objects by mouth; and
6. visual agnosia, an inability to recognize objects or drawings of objects (see Chapter 8).
416 ! CHAPTER 11
–2.0
–1.8
–1.6
–1.4
–1.2
–1.0
–0.8
–0.6
–0.4
–0.2
0
0.2
0.4
0.6
Location of lesion in spinal cord
Sacral
(low)
Lumbar
Anger
Low
thoracic
High
thoracic
Cervical
(high)
Change in emotionality subsequent to spinal-cord lesion
Increase
Spinal cord
Decrease
Fear
Figure 11-23
Losing Emotion Spinal injury reduces
the experience of emotion. The extent
of the loss of emotionality is greatest
when the lesion is high on the spine.
Adapted from Principles of Behavioral
Neuroscience (p. 339), by J. Beatty, 1995,
Dubuque, IA: Brown & Benchmark.
Somatic marker hypothesis. Posits
that “marker” signals arising from
emotions and feelings act to guide
behavior and decision making, usually
in an unconscious process.
Klüver-Bucy syndrome. Behavioral
syndrome, characterized especially by
hypersexuality, that results from bilateral
injury to the temporal lobe.
CH11.qxd 2/3/05 4:20 PM Page 416
ls
le
ll
Visual agnosia is due to damage to the ventral visual stream in the temporal lobe,
but the other symptoms are related to the amygdala damage. The tameness and loss of
fear after amygdalectomy is especially striking. Monkeys that normally show a strong
aversion to stimuli such as snakes show no fear of them whatsoever. In fact, amygdalectomized
monkeys may pick up live snakes and even put them in their mouths.
Although the Klüver-Bucy syndrome is not common in humans, because bilateral
temporal lobectomies are rare, symptoms of the syndrome can be seen in people with
certain forms of encephalitis, a brain infection (see “Meningitis and Encephalitis” on
page 42). In some cases, an encephalitis centered on the base of the brain can damage
both temporal lobes and produce many Klüver-Bucy symptoms, including especially
indiscriminate sexual behavior and the tendency to examine objects by mouth.
The role of the amygdala in Klüver-Bucy syndrome points to its central role in
emotion. So does electrical stimulation of the amygdala,which produces an autonomic
response (such as increased blood pressure and arousal) as well as a feeling of fear. Although
this production of fear by the brain in the absence of an obvious threat may
seem odd, fear is important to a species’ survival. To improve their chances of surviving,
most organisms using fear as a stimulus minimize their contact with dangerous
animals, objects, and places and maximize their contact with things that are safe.
The awareness of danger and safety has both an innate and a learned component,
as Joe LeDoux (1996) emphasized. The innate component,much as in the IRMs, is the
automatic processing of species-relevant sensory information—specifically, sensory
inputs from the visual, auditory, and olfactory systems. The importance of olfactory
inputs is not obvious to humans, whose senses are dominated by vision. But, for other
animals, olfactory cues often predominate, and there is a major input of olfactory information
directly into the amygdala (you can see this connection in the human brain
in Figure 11-19).
Thus, a rat that has never encountered a ferret shows an immediate fear response
to the odor of ferret.Other novel odors (such as peppermint or coffee) do not produce
an innate fear reaction. The innate response triggers an autonomic activation that stimulates
conscious awareness of danger.
In contrast, the learned component of fear consists of the avoidance of specific animals,
places, and objects that the organism has come to associate with danger. The organism
is not born with this avoidance behavior prewired. In a similar way, animals
learn to increase contact with environmental stimuli that they associate with positive
outcomes, such as food or sexual activity or, in the laboratory, drugs. Damage to the
amygdala interferes with all these behaviors. The animal loses not only its innate fears
but also its acquired fears and preferences for certain environmental stimuli.
To summarize, the amygdala is required for species survival. It influences autonomic
and hormonal responses through its connections to the hypothalamus. It influences
our conscious awareness of the positive and negative consequences of events and
objects through its connections to the prefrontal cortex.
The Prefrontal Cortex and Emotional Behavior
At about the same time that Klüver and Bucy began studying their monkeys, Carlyle
Jacobsen was studying the effects of frontal lobotomy on the cognitive capacities of two
chimpanzees. A frontal lobotomy destroys a substantial amount of brain tissue as the
result of inserting a sharp instrument into the frontal lobes and moving it back and
forth.
In 1935, Jacobsen reported that one of the chimps that had been particularly neurotic
before being subjected to this procedure, became more relaxed after it. Incredibly,
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 417
CH11.qxd 2/3/05 4:20 PM Page 417
ls
le
ll
a leading Portuguese neurologist of the time, Egas Moniz, seized on this observation as
a treatment for behavioral disorders in humans, and the frontal lobotomy was initiated
as the first form of psychosurgery. One procedure is illustrated in Figure 11-24.
The use of psychosurgery, which refers to any neurosurgical technique intended to
alter behavior, grew rapidly in the 1950s. In North America alone, nearly 40,000 people
received frontal lobotomies as a treatment for psychiatric disorders. Not until the
1960s was there any systematic research into the effects of frontal lesions on social and
emotional behavior. By this time, the frontal lobotomy had virtually vanished as a
“treatment.” There is now little doubt that prefrontal lesions in various species, including
humans, have severe effects on social and emotional behavior.
Agnes is a case in point. We met Agnes at the psychiatric hospital where we met
Roger, whose indiscriminate eating was described at the beginning of this chapter. At
the time, Agnes, a 57-year-old woman, was visiting one of the nurses. Agnes had, however,
once been a patient.
She had been subjected to a procedure known as a frontal leukotomy because her
husband, an oil tycoon, felt that she was too gregarious. Evidently, he felt that her “loose
lips”were a detriment to his business dealings.He convinced two psychiatrists that she
would benefit from psychosurgery, and her life was changed forever.
To perform a leukotomy, as illustrated in Figure 11-24, a surgeon uses a special
knife called a leukotome to sever the connections of a region of the inferior frontal cortex,
including especially the orbital cortex (see Figure 11-21). The first thing that we
noticed about Agnes was that she exhibited no outward sign of emotion. She had virtually
no facial expression.
In our conversations with her, however, we quickly discovered that she had considerable
insight into the changes brought about by the leukotomy. In particular, she
indicated that she no longer had any feelings about things or most people, although,
curiously, she was attached to her dog. She said that she often just felt empty and much
like a zombie.
Her only moment of real happiness in the 30 years since her operation was the sudden
death of her husband, whom she blamed for ruining her life. Unfortunately, Agnes
had squandered her dead husband’s considerable wealth as a consequence of her inability
to plan or organize. This inability, we have seen, is another symptom of prefrontal
injury.
The orbital region of the inferior prefrontal area has direct connections with the
amygdala and hypothalamus. Stimulation of this area can produce autonomic responses,
and, as we saw in Agnes, damage to this area can produce severe personality
change characterized by apathy and loss of initiative or drive. The orbital cortex is likely
responsible for the conscious awareness of emotional states that are produced by the
rest of the limbic system, especially the amygdala.
Agnes’s loss of facial expression is also fairly typical of frontal-lobe damage. In fact,
people with frontal-lobe injuries or those who suffer from schizophrenia are usually
impaired both at producing and perceiving facial expressions, including a wide range
of expressions found in all human cultures—happiness, sadness, fear, anger, disgust,
and surprise (Kolb & Taylor, 2000). As with J. P.’s agenesis, described earlier, it is difficult
to imagine how such people can function effectively in our highly social world
without being able to recognize the emotions of others.
Although facial expression is a key part of recognizing emotion, so is tone of voice,
or prosody. Frontal-lobe patients are devoid of prosody, both in their own conversations
and in understanding the prosody of others. The lost ability to comprehend or
produce emotional expression in both faces and language partly explains the apathy of
frontal-lobe patients. In some ways, they are similar to spinal-cord patients who have
lost autonomic feedback and so can no longer feel the arousal associated with emotion.
418 ! CHAPTER 11
Psychosurgery. Any neurosurgical
technique intended to alter behavior.
Figure 11-24
Transorbital Leukotomy In this
procedure, a leukotome is inserted
through the bone of the eye socket
and the inferior frontal cortex is
disconnected from the rest of the brain.
Learn more about the history of
the frontal leukotomy and other
psychosurgeries at the Web site at www.
worthpublishers.com/kolb/chapter11
CH11.qxd 2/3/05 4:20 PM Page 418
ls
le
ll
Frontal-lobe patients can no longer either read emotion in other people’s faces and
voices or experience it in their own.
Some psychologists have proposed that our own facial expressions may provide us
with important clues to the emotions that we are feeling. This idea has been demonstrated
in experiments reviewed by Pamela Adelmann and Robert Zajonc (1989). In
one such study, people were required to contract their facial muscles by following instructions
about which parts of the face to move. Unbeknown to the participants, the
movements produced happy and angry expressions. Afterward, they viewed a series of
slides and reported how the slides made them feel.
They said that they felt happier when they were inadvertently making a happy face
and angrier when the face that they were making was one of anger. Frontal-lobe patients
presumably would have no such feedback from their own facial expressions,
which could be a reason why their emotional experiences are dampened.
Emotional Disorders
As detailed in Chapter 7, a highly disruptive emotional disorder is major depression,
characterized by abnormal regulation of feelings of sadness and happiness.A depressed
person feels severely despondent for a prolonged time. Depression is common in our
modern world, with a prevalence of at least 10 percent of the population.
Depression has a genetic component. It not only runs in families but also frequently
tends to be found in both members of a pair of identical twins. The genetic component
in depression implies a biological abnormality, but the cause remains unknown.
The strongest evidence supporting a biological cause of depression comes from the
fact that about 70 percent of depressed people can be treated with one of several antidepressant
drugs. This success rate has made antidepressants among the most widely
prescribed classes of drugs in the world. As summarized in Table 11-2, antidepressants
act on synapses (especially noradrenaline- and serotonin-containing synapses)
by increasing the amount of available transmitter at them. The major projections of
noradrenaline- and serotonin-containing cells to the limbic system imply that the activity
of limbic regions, including the prefrontal cortex, is abnormal in depression (see
Chapter 15).
Excessive anxiety is an even more common emotional problem than depression.
Anxiety disorders, including posttraumatic stress disorder, phobias, and obsessive compulsive
disorder, are estimated to affect from 15 to 35 percent of the population. Symptoms
include persistent fears and worries in the absence of any direct threat, usually
accompanied by various physiological stress reactions, such as rapid heartbeat, nausea,
and breathing difficulty, as described in “Anxiety Disorders” on page 420. As with depression,
the root cause of anxiety disorders is not known, but the effectiveness of drug
treatments implies a biological basis.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 419
Types of Antidepressant Medications
Drug type Action Examples
Tricyclic antidepressants Block reuptake of serotonin Imipramine
and noradrenaline
MAO inhibitors Block activity of monoamine oxidase Iproniazid
Selective serotonin reuptake Block reuptake of serotonin Fluoxetine (Prozac)
inhibitors (SSRIs) Sertraline (Zoloft)
Paroxetine (Paxil)
Table 11-2
CH11.qxd 2/3/05 4:20 PM Page 419
ls
le
ll
The most widely prescribed anxiolytic (antianxiety) drugs, which are detailed in
Chapter 7, are the benzodiazepines, such as Valium, Librium, and Xanax. These
drugs are thought to be effective because of their agonistic action on the GABAA receptor
(see Figure 7-8). Although GABAA receptors are found throughout the brain,
the amygdala has an especially high concentration. The infusion of benzodiazepines
Anxiety Disorders
Focus on Disorders
Animals normally become anxious at times, especially when
they are in obvious danger. But anxiety disorders are different.
They are characterized by intense feelings of fear or anxiety
that are not appropriate for the circumstances. People
with an anxiety disorder have persistent and unrealistic worries
about impending misfortunes. They also tend to suffer
multiple physical symptoms attributable to hyperactivity of
the sympathetic nervous system.
G. B.’s case is a good example. He was a 36-year-old man
with two college degrees who began to experience severe
spells that were initially diagnosed as some type of heart condition.
He would begin to breathe heavily, sweat, experience
heart palpitations, and sometimes suffer pains in his chest and
arms. During these attacks, he was unable to communicate
coherently and would lie helplessly on the floor until an ambulance
arrived to take him to an emergency room.
Extensive medical testing and multiple attacks in a period
of about 2 years eventually led to the diagnosis of generalized
anxiety disorder. Like most of the 5 percent of the U.S. population
who suffer an anxiety disorder at some point in their
lives, G. B. was unaware that he was overly anxious.
The cause of generalized anxiety is difficult to determine,
but one likely explanation is that these attacks are related
to the cumulative effect of general stress. Although G.
B. appeared outwardly calm most of the time, he had been
a prodemocracy activist in communist Poland, a dangerous
position to adopt. Because of the dangers, he and his family
eventually had to escape from Poland to Turkey, and from
there they went to Canada. G. B. may have had continuing
worries about the repercussions of his political activities—
worries (and stress) that eventually found expression in generalized
anxiety attacks.
The most common and least disabling type of anxiety
disorders are phobias. A phobia pertains to a clearly defined
dreaded object (such as spiders or snakes) or some greatly
feared situation (such as enclosed spaces or crowds). Most
people have mild aversions to some types of stimuli.
Such an aversion becomes a phobia only when a person’s
feelings toward a disliked stimulus lead to overwhelming
fear and anxiety. The incidence of disabling phobias is
surprisingly high, being estimated to affect at least one in ten
people. For most people with a phobia, the emotional reaction
can be controlled by avoiding what they dread.
Panic disorder has an estimated incidence on the order
of 3 percent of the population. The symptoms of panic disorder
include recurrent attacks of intense terror that come on
without warning and without any apparent relation to external
circumstances. Panic attacks usually last only a few minutes,
but the experience is always terrifying. There is sudden
activation of the sympathetic nervous system, leading to
sweating, a wildly beating heart, and trembling.
Although panic attacks may occur only occasionally,
the victim’s dread of another episode may be continual.
Consequently, many people with panic disorders also experience
agoraphobia, a fear of public places or situations in
which help might not be available. This phobia makes some
sense because a person with a panic disorder may feel particularly
vulnerable to having an attack in a public place.
Freud believed that anxiety disorders were psychological
in origin and treatable with talking therapies in which
people confronted their fears. But anxiety disorders are now
known to have a clear biological link. These disorders are
most effectively treated with benzodiazepines, of which diazepam
(Valium) is the best known. Alprazolam (Xanax) is
the most commonly prescribed drug for panic attacks. Benzodiazepines
act by augmenting GABA’s inhibitory effect
and are believed to exert a major influence on neurons in the
amygdala.
CH11.qxd 2/3/05 4:20 PM Page 420
ls
le
ll
into the amygdala blocks fear, suggesting that the amygdala may be the site of their
action.
Why would the brain have a mechanism for benzodiazepine action? It certainly did
not evolve to allow us to take Valium. Probably this mechanism is part of a system that
both increases and reduces anxiety levels. The mechanism for raising anxiety seems to
entail a compound known as diazepam-binding inhibitor. This compound appears to
bind antagonistically with the GABAA receptor, resulting in greater anxiety.
There are times when such an increase in anxiety is beneficial, especially if we are
drowsy and need to be alert to deal with some kind of crisis. Impairment of this mechanism
or the one that reduces anxiety can cause serious emotional problems, even anxiety
disorders (Chapter 15).
CONTROL OF REGULATORY BEHAVIOR
Feeding behaviors are central to our existence. After all, we must eat and drink to live.
But there is more to feeding behavior than sustenance alone.We also derive great pleasure
from eating and drinking.
In fact, for many people, eating is a focus of daily life, if not for survival, for its centrality
to social activities, from get-togethers with family and friends to business meetings
and even to identification with a group. Are you a gourmet, a vegetarian, or a
snack-food junkie? Do you diet? In this section, we focus mainly on the control of eating
in humans, but we also consider how homeostatic mechanisms control our intake
of fluids.
Controlling Eating
Control over eating is a source of frustration and even grief for many people in the developed
world. In 2000, the World Health Organization identified obesity, the excessive
accumulation of body fat, as a worldwide epidemic. The United States is a case in
point. From 1990 to 2000, the percentage of overweight people has increased from
about 50 percent to 60 percent of the population. The proportion of people considered
obese has increased from about 12 percent to 20 percent.
The increasing numbers of overweight and obese children and adults persist despite
a substantial decrease in fat intake in American diets.What behaviors might cause
persistent weight gain? One key to understanding weight gain in the developed world
is evolutionary.
In Review .
Emotion and motivation have common autonomic responses (sweating and rapid heartbeat),
subjective feelings (fear or trust, joy or pain), and a cognitive component—what we
think about the arousing situation. The autonomic responses result from the activity of the
hypothalamus and related structures. Psychologists have proposed that, when the body experiences
an autonomic reaction and intense feelings, the brain creates a story to explain
those experiences. The amygdala and the orbitofrontal-cortex circuits contribute to our
feelings and our motives. Our emotional thoughts and thinking that motivates us both
likely result from activity throughout the cerebral hemispheres. Abnormalities in the neural
circuits that produce emotional and motivated behavior are responsible for society’s
most pervasive behavioral disorders—the anxiety disorders and depression.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 421
Generalized anxiety disorder.
Persistently high levels of anxiety often
accompanied by maladaptive behaviors
to reduce anxiety; thought to be caused
by chronic stress.
Phobia. Clearly defined dreaded object
or greatly feared situation.
Panic disorder. Recurrent attacks of
intense terror that come on without
warning and without any apparent
relation to external circumstances.
Obesity. Excessive accumulation of
body fat.
CH11.qxd 2/3/05 4:20 PM Page 421
ls
le
ll
422 ! CHAPTER 11
Even 40 years ago, much of our food was only seasonally available. In a world
with uncertain food availability, it makes sense to store excess body calories in the
form of fat to be used later when food is scarce. Down through history and in many
cultures today, plumpness is desirable as a standard of beauty and a sign of health
and wealth.
In a postindustrial society where food is continuously and easily available, as it is
today, being overweight may not be the healthiest strategy. People eat as though food
will be scarce and fail to burn off the extra calories by exercising, and the result is apparent.
About half of the U.S. population has dieted at some point in their lives. At any
given time, at least 25 percent report that they are currently on a diet. For a comparison
of how some well known diets perform, see “Weight-Loss Strategies.”
Eating disorders entail being either underweight or overweight. Most Americans
are overweight but live in a culture obsessed with slimness. The human control system
for feeding has multiple neurobiological inputs, including cognitive factors such as
Weight-Loss Strategies
Focus on Disorders
Among the wide range of diets and weight-loss strategies on
the market, none have stopped the obesity epidemic. Diets
range widely in their recommended proportions and types
of fats, carbohydrates, and proteins allowed. In the past
few years, low-carbohydrate diets have become popular—
Atkins, Zone, and South Beach among them.
Proponents of these plans claim significant weight
loss and good health, without hunger. The accompanying
charts profile the dietary proportions recommended for
the beginning phase of the Atkins plan, for the Zone diet,
and for a diet high in nonsaturated fats and low in carbohydrates.
Shown for comparison is the dietary compostition
that the American Heart Association recommended
in 2000.
What scientific evidence supports the claims made by
these diets? Are they fads or do they really work? The difficulty
in reaching scientific conclusions about a diet’s effectiveness
is that studies need random samples of individuals
on different diets to compare over a period of at least 1 year.
Few such studies have been designed, and the conclusion
of a recent review by Arne Astrup and colleagues (2004) is
that weight loss is associated with the restriction of energy
intake and the duration of the diet but not with carbohydrate
restriction.
Adapted from Astrup et al., 2004.
Carbohydrate Nonsaturated fat Saturated fat Protein
Atkins diet,
(induction phase) Zone diet Willett diet
American Heart Association
(year 2000)
15%
10%
20%
55%
15%
10%
30%
45%
30%
40%
20%
10%
5%
36%
23%
36%
CH11.qxd 2/3/05 4:20 PM Page 422
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 423
Link to the Web site at www.worth
publishers.com/kolb/chapter11 to
learn more about eating disorders.
thoughts about food. These cognitive factors also include the association between
environmental cues (e.g., watching television or studying) and the act of eating. The
constant pairing of such cues with eating can result in the cues alone becoming a motivation,
or incentive, to eat. We return to this phenomenon in the discussion of rewards
and addictions at the end of the chapter.
Anorexia nervosa is a disorder with a huge cognitive component—namely, selfimage.
Anorexia is especially identified with adolescent girls. A person’s body image is
highly distorted in anorexia. This misperception leads to an exaggerated concern with
being overweight, spiraling to excessive dieting, compulsive exercising, and severe, potentially
life threatening weight loss.
The neurobiological control of feeding behavior in humans is not as simple as it is
in the fly described earlier in the chapter. The multiple inputs to the human control
system for feeding come from three major sources: the cognitive factors already introduced,
the hypothalamus, and the digestive system.
For example, in one study, 63 participants followed
either of two diets in which the proportions of nutrients
are essentially reversed (Foster et al., 2003). In the lowcarbohydrate–
high-protein Atkins plan, the percentage of
carbohydrates allowed ranges from 5 percent in the induction
phase to a maximum of 19 percent in the maintenance
phases. Protein is at least 36 percent and fat as much as 59
percent of the menu, depending on the phase. In an energyrestricted
diet that is high in carbohydrates and low in protein,
the allowable percentages are 60 percent carbohydrate
and only 15 percent protein and 25 percent fat. After 6
months, the low-carbohydrate group on Atkins had lost a
larger percentage of body weight on average (7 percent versus
3 percent); but, after 1 year, the difference was no longer
significant (4.4 percent versus 2.5 percent).
Thus, the low-carbohydrate diet was superior in the
short run but not in the long run, a finding supported by
those of two other studies reviewed by Astrup’s group. Two
problems with all studies are that drop-out rates tend to be
high and it is difficult to ensure compliance with the diets.
Therefore, to date, there is no evidence that, in the long
term, the low-carbohydrate diets are superior.
What mechanisms might cause weight loss? The greater
weight loss in the short term on the low-carbohydrate diets
appears to be a result of increased loss of stored fat. There is
no evidence that any diet affects the expenditure of energy,
a claim made by the Atkins diet. Crankiness due to low carbohydrate
intake, which lowers serotonin levels, has been
reported.
Another possible explanation, but not proved, for the
short-term success of the low-carbohydrate diet is the combination
of restricted food choices and enhanced feeling of
satiety produced by its high protein content. The drop-out
rate may be related to a long-term dissatisfaction with the restricted
food choices, but that possibility, too, needs to be
confirmed. Astrup concludes that, at present, the only certain
solution to weight loss appears to be a permanent switch
to a diet reduced in calories and fat combined with increased
physical activity. One of the largest sources of calories
is sugar (a carbohydrate) in highly refined foods.
Finally, although not specifically related to dieting, there
is increasing evidence of the health-protective effects of diets
that are high in fruits, vegetables, legumes, and whole grains
and that include fish, nuts, and low-fat dairy products. Perhaps
the best known of such diets is the Mediterranean diet,
which has well-documented benefits in reducing the risk of
heart disease as well as obesity (Panagiotakos et al., 2004).
Such diets need not be restricted in fat intake as long as people
are active and energy intake does not exceed energy
expenditure. Although there has been some debate about
whether the effects of such diets on health are partly biased
by genetics or culture, systematic clinical trials of people at
risk for cardiovascular disease elsewhere in the world have
shown clear reductions in cardivascular disease and death.
Anorexia nervosa. Exaggerated
concern with being overweight that leads
to inadequate food intake and often
excessive exercising; can lead to severe
weight loss and even starvation.
CH11.qxd 2/3/05 4:20 PM Page 423
ls
le
ll
THE DIGESTIVE SYSTEM AND THE CONTROL OF EATING
The digestive tract, illustrated in Figure 11-25, begins in the mouth and ends at the
anus. Food travels from the oral cavity to the stomach through the esophagus. The
stomach, which is a storage reservoir, secretes hydrochloric acid, which starts to break
food into smaller particles, and pepsin, an enzyme that breaks proteins down into
amino acids.
The partly broken down food then moves to the upper part of the small intestine
through the duodenum, where digestive enzymes produced in the gall bladder and pancreas
further break the food down to allow the absorption of amino acids, fats, and simple
sugars into the bloodstream.Most of the remaining water and electrolytes in food are
absorbed by the large intestine, and the waste passes out of the body through the anus.
The digestive system extracts three types of nutrients for the body: lipids (fats),
amino acids (the building blocks of proteins), and glucose (sugar). Each is a specialized
form of energy reserve. Because we require varying amounts of these reserves, depending
on what we are doing, the body has detector cells to keep track of the level of
each nutrient in the bloodstream.
Glucose is the body’s primary fuel and is virtually the only energy source for the
brain. Because the brain requires glucose even when the digestive tract is empty, the
liver acts as a short-term reservoir of glycogen, a starch that acts as an inert form of
glucose. When blood-sugar levels fall, as when we are sleeping, detector cells tell the
liver to release glucose by converting glycogen into glucose.
Thus the digestive system functions mainly to break down food, and the body
needs to be apprised of how well this breakdown is proceeding. Feedback mechanisms
provide such information.When food reaches the intestines, it interacts with receptors
there to trigger the release of at least 10 different peptide hormones, including one
known as cholecystokinin (CCK).
The released peptides inform the brain (and perhaps other organs in the digestive
system) about the nature and quality of the food in the gastrointestinal tract.The level of
CCK appears to play a role in satiety, or the feeling of having eaten enough. For example,
if CCK is infused into the hypothalamus of an animal, the animal’s appetite diminishes.
THE HYPOTHALAMUS AND THE CONTROL OF EATING
Feeding behavior is influenced by hormones including insulin, growth hormone, and
sex steroids. These hormones stimulate and inhibit feeding, and they aid in the conversion
of nutrients into fat and the conversion of fat into glucose. Not surprisingly, the
hypothalamus, which controls hormone systems, is the key brain structure in feeding.
Investigation into the role of the hypothalamus in the control of feeding began in
the early 1950s, when researchers discovered that damage to the lateral hypothalamus
in rats caused the animals to stop eating, a symptom known as aphagia (from the
Latin a, meaning “not,” and phagia, meaning “chew”). In contrast, damage to the
ventromedial hypothalamus (VMH), caused the animals to overeat, a symptom known
as hyperphagia. A VMH-lesioned rat that overate to the point of obesity is shown in
the Procedure section of Experiment 11-1. The Results section reveals that the VMHlesioned
rat weighed more than a kilogram, three times the weight of her normal sister,
which was 340 grams.
At about the same time, researchers also found that electrical stimulation of the
lateral hypothalamus elicits feeding, whereas stimulation of the ventromedial hypothalamus
inhibits feeding. The opposing effects of injury and stimulation to these two
hypothalamic regions led to the idea that the lateral hypothalamus signals “eating on,”
whereas the VMH signals “eating off.” This model quickly proved to be too simple,
however.
The lateral hypothalamus contains not only cell bodies but also fiber bundles passing
through it, and damage to either can produce aphagia. Similarly, damage to fibers
424 ! CHAPTER 11
Liver
Stomach
Pancreas
Gallbladder
Duodenum
Large
intestine
Small
intestine
Anal canal
Rectum
Oral
cavity
Tongue
Esophagus
Figure 11-25
The Digestive System
Aphagia. Failure to eat; may be due to
an unwillingness to eat or to motor
difficulties, especially with swallowing.
Hyperphagia. Disorder in which an
animal overeats, leading to significant
weight gain.
CH11.qxd 2/3/05 4:20 PM Page 424
ls
le
ll
passing through the VMH often causes injury as well to the paraventricular
nucleus of the hypothalamus.And damage to the paraventricular
nucleus alone is now known to produce hyperphagia. Clearly,
then, there is more to the hypothalamus’s role in the control of feeding
than the activities of the lateral and ventromedial hypothalamus alone.
In the half-century since the first studies on the hypothalamus’s
role in feeding, researchers have learned that damage to the lateral
and ventromedial hypothalamus and to the paraventricular nucleus
has multiple effects. These effects include changes in hormone levels
(especially that of insulin), in sensory reactivity (the taste and attractiveness
of food is altered), in glucose and lipid levels in the blood,
and in metabolic rate. The general role of the hypothalamus is to act
as a sensor for the levels of lipids, glucose, hormones, and various
peptides. For example, groups of hypothalamic neurons sense the
level of glucose (so-called glucostatic neurons) as well as the level of
lipids (so-called lipostatic neurons).
The sum of the activity of all such hypothalamic neurons creates
a very complex homeostat controlling feeding. Figure 11-26 shows
that this homeostat receives inputs from three sources: from the digestive
system (such as information about blood-glucose levels),
from hormone systems (such as information about the level of CCK),
and from parts of the brain that process cognitive factors.We turn to
these cognitive factors next.
COGNITIVE FACTORS AND THE CONTROL
OF EATING
Cognitive factors are especially important for the control of eating in
humans. Just thinking about a favorite food is often enough to make
us feel hungry. The cognitive aspect to feeding includes not only the
images of food that we pull from memory but also external sensations,
especially food-related sights and smells. In addition, learned
associations, such as learned taste aversions discussed earlier, are related
to feeding.
The neural control of cognitive factors probably originates in
multiple brain regions. Two structures are clearly important: the
amygdala and the inferior prefrontal cortex.Damage to the amygdala
alters food preferences and abolishes taste-aversion learning. These
effects are probably related to the amygdala’s efferent connections to
the hypothalamus.
The amygdala’s role in regulating species-typical behaviors is
well established, but the role of the inferior prefrontal cortex is more
Procedure
Rat Intact brain of sister rat brain with lesion
The VMH plays a role in controlling the cessation of eating.
Damage to the VMH results in prolonged and dramatic
weight gain.
Conclusion
Question: Does the hypothalamus play a role in eating?
EXPERIMENT 11-1
The ventromedial
hypothalamus (VMH)
of the rat on the
right was damaged,
and her body weight
was monitored for a
year. Her sister on the
left is normal.
The VMH-lesioned rat showed a dramatic
increase in food intake and body weight.
0 1
Time (months)
Lesioned rat
Control rat
2 3 4 5 6 7 8 9 10 11 12
500
1000
1500
100
Body weight (gm)
Results:
Hypothalamus
Digestive
system
Cognitive
factors
Hormones
Sensory
systems
Prefrontal
cortex
Eating
Amygdala
Figure 11-26
Simple Model of Control of Feeding
Behavior
CH11.qxd 2/3/05 4:20 PM Page 425
ls
le
ll
426 ! CHAPTER 11
difficult to pin down.Rats and monkeys with damage to the orbital cortex lose weight, in
part because they eat less.Humans with orbital injuries are invariably slim, but we know
of no formal studies on their eating habits.The inferior prefrontal cortex receives projections
from the olfactory bulb, and cells in this region do respond to smells.Because odors
influence the taste of foods, damage to the inferior prefrontal cortex likely decreases eating
because of diminished sensory responses to food odor and perhaps to taste.
An additional cognitive factor in the control of eating is the pleasure that we derive
from it, especially from eating foods with certain tastes, such as chocolate. What
pleasure is and how the brain produces it are topics discussed at the end of this chapter
in the context of reward. At this point, simply keep in mind that pleasure and its
absence are cognitive factors in the control of eating.
Finally, Randy Seeley and Stephen Woods (2003) have noted that, in spite of the
problem that people now appear to have with weight gain, adult mammals do a masterful
job of matching their caloric intake to caloric expenditure. Consider that a typical
man eats 900,000 calories per year. To gain just one extra pound requires him to eat
4000 calories more than are burned in that year. This increase amounts to only 11 calories
per day, or one potato chip.According to Seeley and Woods, the average weight gain
in the U.S population is less than 1 pound per year. This weight gain corresponds to an
error in homeostasis of less than 0.5 percent, which is a rather small error. Seeley and
Woods conjectured that the nervous sytem must juggle two competing challenges in
weight control. The first is to maintain adequate stores of fuel and the second is to provide
fuel for current cellular functions. They noted that the current obesity “epidemic”
could be the result of factors that alter the sensing of stored fuel, the sensing of ongoing
fuel availability, or the integration of these two types of signal. Treatment strategies for
obesity will require an understanding of what these factors are and how they operate.
Controlling Drinking
About 70 percent of the human body is composed of water that contains a range of
chemicals that participate in the hundreds of chemical reactions of bodily functions.
Homeostatic mechanisms control water levels (and hence chemical concentrations)
within rather narrow limits. These mechanisms are essential because the rate of a
chemical reaction is partly determined by how concentrated the supplies of participating
chemicals are.
As with eating, we drink for many reasons.We consume some beverages, such as
coffee, wine, beer, and juice, as part of social activities or just because they taste good.
We drink water for its health benefits, to help wash down a meal or to intensify the flavor
of dry foods. On a hot day, we drink water because we are thirsty, presumably because
we have lost significant moisture through sweating and evaporation.
Although we think of thirst as a single phenomenon, there are actually two kinds
of thirst. Osmotic thirst results from an increase in the concentrations of chemicals,
known as solutes, in the body fluids. Hypovolemic thirst results from a loss of overall
fluid volume from the body. Here we consider each kind briefly.
OSMOTIC THIRST
The solutes found inside and outside cells in the body have an ideal concentration for
the body’s chemical reactions. This concentration requires a kind of thermostat,much
like the one for body temperature. Deviations from the ideal solute concentration activate
systems to reestablish that concentration.
Consider what happens when we eat salty foods, such as potato chips. The salt
(NaCl) spreads through the blood and enters the extracellular fluid that fills the spaces
Osmotic thirst. Results from an
increased concentration of chemicals,
known as solutes, in body fluids.
Hypovolumic thirst. Produced by a
loss of overall fluid volume from the
body.
CH11.qxd 2/3/05 4:20 PM Page 426
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 427
between our cells. This produces a shift away from the ideal solute concentration. Receptors
in the hypothalamus along the third ventricle detect the altered solute concentration
and relay this message to various hypothalamic areas that, in turn, stimulate us
to drink in response to osmotic thirst. In addition, other messages are sent to the kidneys
to reduce water excretion.
HYPOVOLEMIC THIRST
Unlike osmotic thirst, hypovolemic thirst arises when the total volume of body fluids
declines,motivating us to drink more and replenish their supplies. In contrast with osmotic
thirst, however, hypovolemic thirst encourages us to choose something other
than water, because water would dilute the solute concentration in the blood. Rather,
we prefer to drink flavored beverages that contain salts and other nutrients.
Hypovolemic thirst and its satiation are controlled by a different hypothalamic circuit
from the one that controls osmotic thirst.When fluid volume drops, the kidneys
send a hormone signal (angiotensin) that stimulates midline hypothalamic neurons.
These neurons, in turn, stimulate drinking.
CONTROL OF NONREGULATORY BEHAVIOR
Unlike feeding, which organisms must repeatedly do to survive, sexual behavior—
notwithstanding procreation, which is essential to the survival of the species—is not
essential for an individual organism’s survival. Yet sexual activity is of enormous psychological
importance to humans. Sexual themes repeatedly appear in our art, literature,
and films. They also bombard us in the advertising that we create to help sell
products.
Indeed, in Sigmund Freud’s psychodynamic theory of human behavior, sexual
drives are central. Such significance makes it all the more important to understand how
human sexual behavior is controlled. The answer lies in both gonadal hormones and
brain circuits.
Effects of Sex Hormones on the Brain
In Chapter 6, we encountered the influence of gonadal hormones on the brain when
we considered how a male’s Y chromosome controls the differentiation of embryonic
gonad tissue into testes, which in turn secrete testosterone. Testosterone masculinizes
both the sex organs and the brain during development. This process is an organizing
In Review .
Feeding is a regulatory behavior that has a large cognitive component in humans. The
hypothalamus is the principal brain structure in the control of eating. Three of its regions—the
lateral hypothalamus, the ventromedial hypothalamus, and the paraventricular nucleus—
play especially important roles. Groups of neurons in the hypothalamus act as sensors to detect
the levels of glucose, lipids, and peptides in the blood. Neural control of the cognitive
factors in eating probably includes multiple brain regions. The brain also motivates us to drink
whenever solutes in the blood deviate from ideal levels or whenever there is a significant drop
in the body’s volume of fluids. In either case, receptors detect the shifts, and neurons in the
hypothalamus stimulate the experience of thirst.
CH11.qxd 2/3/05 4:20 PM Page 427
ls
le
ll
428 ! CHAPTER 11
effect of gonadal hormones. A major organizing effect that gonadal hormones have on
the brain is in the hypothalamus, especially the preoptic area of the medial hypothalamus.
But organizing effects also operate in other nervous system regions, notably the
amygdala, the prefrontal cortex, and the spinal cord.
These sex-related differences in the nervous system make sense behaviorally. After
all, animal courtship rituals differ between the sexes, as do copulatory behaviors, with
females engaging in sexually receptive responses and males in mounting ones. The production
of these sex differences in behaviors depends on the action of gonadal hormones
on the brain both during development and during adulthood.
The actions of hormones on the adult brain are referred to as activating effects, in
contrast with the developmental organizing effects. Here we consider organizing and
activating effects separately.
ORGANIZING EFFECTS OF SEX HORMONES
During fetal development, as you know, a male’s testes produce male hormones, the
androgens. In the developing rat (the species in which the organizing effects of gonadal
hormones have been most extensively studied), androgens are produced during the last
week of fetal development and the first week after birth. The androgens produced at
this time greatly alter both neural structures and later behavior. For example, the hypothalamus
and prefrontal cortex of a male rat differ structurally both from those of
female rats and from those of males that were not exposed to androgens during their
development.
Furthermore, in adulthood,males with little exposure to the androgen testosterone
during development behave like genetic female rats. If given estrogen and progesterone,
they become sexually “receptive” and display typical female behaviors when
mounted by males. Male rats that are castrated in adulthood do not act in this way.
Sexual dimorphism, the differential development of brain areas in the two sexes
(see Chapter 6), arises from a complex series of steps. Cells in the brain produce aromatase,
an enzyme that converts testosterone into estradiol, one of the female sex hormones
called estrogens. Therefore a female hormone, estradiol, actually masculinizes
a male brain.
Females are not masculinized by the presence of estrogens, because the fetuses of
both sexes produce a liver enzyme (alpha fetoprotein) that binds to estrogen, rendering
it incapable of entering neurons. Testosterone is unaffected by alpha fetoprotein, and
so it enters neurons and is converted into estradiol.
The organizing effects of testosterone are clearly illustrated in the preoptic area of
the hypothalamus, which plays a critical role in the copulatory behavior of male rats.
Comparing this area in males and females, Roger Gorski and his colleagues found a nucleus
about five times as large in the males as in the females (Gorski, 1984). Significantly,
the sexual dimorphism of the preoptic area can be altered by manipulating
gonadal hormones during development. The castration of male rats at birth leads to a
smaller preoptic area, whereas treating infant females with testosterone increases the
preoptic area’s size.
The organizing effects of gonadal hormones are more difficult to study in humans.
The work of John Money and Anke Ehrhardt (1972), however, revealed an important
role of these hormones in human development (see “Androgen-Insensitivity Syndrome
and the Androgenital Syndrome”).
ACTIVATING EFFECTS OF SEX HORMONES
The sexual behavior of both males and females also depends on the actions that gonadal
hormones have on the adult brain. In most vertebrate species, female sexual behavior
varies in the course of an estrous cycle during which the levels of hormones
that the ovaries produce fluctuate. The rat’s estrous cycle is about 4 days long, with
OH
Testosterone
O
CH3
CH3
OH
Estradiol
HO
CH3
Sexual dimorphism. Process whereby
gonadal hormones act on the brain to
produce a distinctly female or male brain.
CH11.qxd 2/3/05 4:20 PM Page 428
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 429
sexual receptivity being only in the few hours during which the production of the
ovarian hormones estrogen and progesterone peaks. These ovarian hormones alter
brain activity, which in turn alters behavior. Furthermore, in female rats, various
chemicals are released after mating, and these chemicals inhibit further mating
behavior.
The activating effect of ovarian hormones can be seen clearly in cells of the hippocampus.
Figure 11-27 compares hippocampal pyramidal neurons taken from female
rats at two points in the estrous cycle: one when estrogen levels are high and the other
when they are low. When estrogen levels are high, more dendritic spines and, presumably,
more synapses emerge. These neural differences during the estrous cycle are
all the more remarkable when we consider that cells in the female hippocampus are
continually changing their connections to other cells every 4 days throughout the animal’s
adulthood.
Androgen-Insensitivity Syndrome and the Androgenital Syndrome
Focus on Disorders
After the testes of a male fetus have formed,
sexual development depends on the actions
of testicular hormones. This dependence
is made extremely clear by studying
people with androgen-insensitivity syndrome.
In this syndrome, an XY (male) fetus
produces androgens, but the body is not
able to respond to them.
Such a genetic male therefore develops
a female appearance, or phenotype, as
shown in the top photograph. Because
their estrogen receptors are not affected by
the syndrome, these people are still responsive
to estrogen produced both by the
adrenal gland and by the testes. As a result,
they develop female secondary sexual
characteristics during puberty, even without
additional hormone treatment. A person
with androgen-insensitivity syndrome
is therefore a genetic male who appears to
be female.
If no Y chromosome is present to induce
the growth of testes, a fetus develops ovaries
and becomes a female. In some cases, however,
the female fetus is exposed to androgens,
producing a syndrome known both as
congenital adrenal hyperplasia and as the
androgenital syndrome. This exposure to androgens
can occur if the adrenal glands of either
the mother or the infant produce an
excessive amount of androgens.
The effects vary, depending on when
the androgens are produced and how
much exposure there is. In extreme cases,
an enlarged clitoris develops that can be
mistaken for a small penis, as shown in the
bottom photograph. In less severe cases,
there is no gross abnormality in genital
structure, but there is a behavioral effect:
these girls show a high degree of tomboyishness.
In early childhood, they identify
with boys and prefer boys’ clothes, toys,
and games. O ne explanation for this behavioral
effect is that the developing brain
is masculinized, thus changing later behavior.
(Top) In androgen-insensitivity syndrome, a
genetic male (XY) is insensitive to gonadally
produced androgens but remains sensitive to
estrogens, leading to the development of a
female phenotype. (Bottom) In congenital
adrenal hyperplasia, a genetic female (XX) is
exposed to androgens produced by the adrenal
gland embryonically, leading to the partial
development of male external genitalia.
Reprinted from Man & Woman, Boy & Girl
by John Money and Anke A. Ehrhardt Reprinted from Man & Woman, Boy & Girl by John Money and Anke A. Ehrhardt
CH11.qxd 2/3/05 4:20 PM Page 429
ls
le
ll
430 ! CHAPTER 11
In males, testosterone activates sexual behavior in two distinctly different ways.
First, the actions of testosterone on the amygdala are related to the motivation to seek
sexual activity. Second, the actions of testosterone on the hypothalamus are needed to
produce copulatory behavior. In the next section, we look at both processes.
The Hypothalamus, the Amygdala,
and Sexual Behavior
The hypothalamus is the critical structure controlling copulatory behaviors in both
males and females. The ventromedial hypothalamus controls the female mating posture,
which in quadrapedal animals is called lordosis and consists of an arching of the
back and an elevation of the rump while the female otherwise remains quite still.Damage
to the VMH abolishes lordosis. The role of the VMH is probably twofold: it controls
the neural circuit that produces lordosis, and it influences hormonal changes in
the female during coitus.
In males, the neural control of sexual behavior is somewhat more complex. The
medial preoptic area, which is larger in males than in females, controls copulation.
Damage to the medial preoptic area greatly disrupts mating performance, whereas
electrical stimulation of this area activates mating, provided that testosterone is circulating
in the bloodstream. Curiously, however, although destruction of the medial preoptic
area stops males from mating, they continue to show interest in receptive females.
For instance, monkeys with lesions in the medial preoptic area will not mate with receptive
females, but they will masturbate while watching them from across the room.
Barry Everitt (1990) studied this phenomenon in an ingenious way. He designed an
apparatus, shown in Figure 11-28, that allowed male rats to press a bar to deliver receptive
females. After males were trained in the use of this apparatus, lesions were made in their
medial preoptic areas. Immediately, their sexual behavior changed.They would still
press the bar to obtain access to females, but they would no longer mate with them.
Apparently, the medial preoptic area controls mating, but it does not control
sexual motivation.The brain structure controlling sexual motivation appears to be
the amygdala.When Everitt trained male rats in his apparatus and then lesioned
their amygdalae, they would no longer press the bar to gain access to receptive females,
but they would mate with receptive females that were provided to them.
In summary, the hypothalamus controls copulatory behavior in both males
and females. In males, the amygdala influences sexual motivation, and it may do
the same among females of species in which sexual activity is not tied to fluctuations
in ovarian hormones, which includes the human species. In other words,
it is likely that the amygdala plays a key role in sexual motivation for human females
as well as for males.
Rat brain
Hippocampus
Pyramidal
neuron
Estrogen
levels high
Estrogen
levels low
Figure 11-27
Hormonal Effects
A comparison of the dendrites of
hippocampal pyramidal neurons at
high and low levels of estrogen in the
rat’s (4-day) estrous cycle reveals that
there are many fewer dendritic spines
in the low period. Adapted from
“Naturally Occurring Fluctuation in Dendritic
Spine Density on Adult Hippocampal Pyramidal
Neurons,” by C. S. Woolley, E. Gould, M.
Frankfurt, and B. McEwen, 1990, Journal of
Neuroscience, 10, p. 1289.
Lordosis
Figure 11-28
Studying Sexual Motivation and
Mating In this experiment, a male rat is
required to press the bar 10 times to gain
access to a receptive female who “drops
in” through a trap door. The copulatory
behavior of the male rat illustrates
mating behavior, whereas the bar
pressing for access to a female rat
illustrates sexual motivation. Adapted from
“Sexual Motivation: A Neural and Behavioral
Analysis of the Mechanisms Underlying
Appetitive and Copulatory Responses of Male
Rats,” by B. J. Everitt, 1990, Neuroscience and
Biobehavioral Reviews, 14, p. 227.
Courtesy of Barry J. Everitt
CH11.qxd 2/3/05 4:20 PM Page 430
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 431
Sexual Orientation, Sexual Identity,
and Brain Organization
An interesting question about human sexual behavior has to do with sexual orientation
—a person’s sexual attraction to the opposite sex or to the same sex. Does sexual orientation
have a neural basis? Although research to answer this question has been limited in
scope, it now appears that differences in the structure of the hypothalamus may form a
basis not only for sexual orientation but also for sexual identity—a person’s feeling either
male or female.
Like rats, humans have sex-related differences in the structure of the hypothalamus.
The preoptic area of male humans can have twice as many neurons as does that
of females, and a region known as the bed nucleus of the stria terminalis is 2.5 times
as large in males (Swaab & Hofman, 1995). Similarly, a hypothalamic region known as
INAH3 is two times as large in males, and a region known as the suprachiasmatic nucleus
(SCN) contains twice as many cells in males as in females.
One hypothesis is that the hypothalamus of homosexual men should be more similar
to the norm for females than for males. This hypothesis turns out to be incorrect,
however. First, no difference between heterosexual and homosexual men is observed in
the size of the preoptic area. Second, the SCN is twice as large in homosexual men as
in heterosexual men.
Some evidence suggests a role for the SCN in sexual behavior in both male and
female rats, and there is strong evidence that manipulating gonadal hormones alters
the structure of the SCN. In contrast with the larger SCN in homosexual men, the
INAH3 is twice as large in the heterosexual brain as in the homosexual brain. These
findings suggest that homosexual men form, in effect, a “third sex” because their hypothalami
differ from those of either females or heterosexual males (Swaab & Hofman,
1995).
No research has been published on the hypothalami of homosexual women. Paul
Vasey (2002) described homosexual behavior in Japanese macaque monkeys. He compared
the hypothalami of this species to data collected in purely heterosexual monkey
species. To date, he has found no unique pattern of hypothalamic organization in homosexual
female Japanese macaques, but we must wonder, Does the homosexual behavior
of these monkeys and human females have a similar basis?
In contrast with homosexuals, transsexuals feel strongly that they have been born
the wrong sex. Their desire to be the opposite sex can be so strong that they undergo
sex-change surgery. Little is known about the causes of transsexuality, but it is generally
assumed to result from a disturbed interaction between brain development and
circulating hormones.
Swaab and Hofman (1995) found that the bed nucleus of the stria terminalis was
female sized in a small group of five male-to-female transsexuals. This finding suggests
the possibility of a biological basis for transsexuality. We must, however, be wary of
drawing cause-and-effect conclusions, especially in such a small sample of people.
If differences in brain organization do exist in people with nontraditional sexual
orientations and sexual identities, what might give rise to these brain differences? Dean
Hamer and his colleagues (1993) studied the incidence of homosexuality in the families
of 114 homosexual men. They recorded a higher-than-average incidence of male
homosexuality on the maternal side of the men’s families but not on the paternal side.
This maternal–paternal difference is most easily explained if a gene on the mother’s
X chromosome is implicated. Further investigation revealed that a large percentage of
homosexual brothers had in common one small area at the tip of the X chromosome
(known as area Xq28). This finding suggests that at least one subtype of male sexual orientation
may be genetically influenced.
Sexual orientation. A person’s sexual
attraction either to the opposite sex or to
the same sex.
Sexual identity. A person’s feeling
either male or female.
CH11.qxd 2/3/05 4:20 PM Page 431
ls
le
ll
We must be cautious in drawing this conclusion, however.William Byne (1994) argued
that, even if certain configurations of genes and neurotransmitters are correlated
with homosexuality, correlation does not prove causation. After all, genes specify proteins,
not sexual behavior.
Conceivably, particular sequences of DNA might cause the brain to be wired
in ways that lead to a particular sexual orientation. But it is equally possible that these
genes could influence the development of certain personality traits that in turn influence
the way in which social experiences contribute to learning a certain sexual
orientation. Clearly, establishing the cause-and-effect connections is not an easy
task.
Cognitive Influences on Sexual Behavior
People think about “sex.” People dream about sex. People make plans about sex. These
behaviors may include activity in the amygdala or the hypothalamus, but they must
certainly also include the cortex. This is not to say that the cortex is essential for sexual
motivation and copulation.
In studies of rats whose entire cortices have been removed, both males and females
still engage in sexual activity, although the males are somewhat clumsy. Nevertheless,
the cortex must play a role in certain aspects of sexual behavior. For instance, imagery
about sexual activity must include activity in the ventral visual pathway of the cortex.
And thinking about sexual activity and planning for it must require the participation
of the frontal lobes.
As you might expect, these aspects of sexual behavior are not easily studied in rats,
and they remain uncharted waters in research on humans.However, changes in the sexual
behavior of people with frontal-lobe injury are well documented. And recall J. P.’s
case, described in “Agenesis of the Frontal Lobes” on page 414.
Although J. P. displayed a loss of inhibition about sexual behavior, frontal-lobe
damage is just as likely to produce a loss of libido (sexual interest). The wife of a man
who, 5 years earlier, had a small tumor removed from the medial frontal region, complained
that she and her husband had since had no sexual contact whatsoever. He was
simply not interested, even though they were both still in their 20s.
The husband said that he no longer had sexual fantasies or sexual dreams and, although
he still loved his wife, he did not have any sexual urges toward her or anyone
else. Such cases clearly indicate that the human cortex has an important role in controlling
sexual behaviors. The exact nature of that role is still poorly understood.
In Review .
Sexual behavior is controlled by a combination of gonadal hormones, neurons in the hypothalamus
and limbic system, and cognitive factors. The hypothalamus controls the details
of copulation in both males and females, whereas the motivation for sexual behavior
is controlled by the amygdala. In contrast with a regulatory behavior such as feeding, nonregulatory
neural control of sexual behavior is affected by the organizing effects of hormones
in the course of development and by their activating effects in adulthood. These
hormones influence the size of subregions and the structure of cells in the hypothalamus,
as well as in the cerebral hemispheres. These anatomical differences presumably account
for some of the differences in sexual behavior, orientation, and identity between males
and females and among individual organisms.
432 ! CHAPTER 11
CH11.qxd 2/3/05 4:20 PM Page 432
ls
le
ll
REWARD
Survival for most animals depends on minimizing contact with certain stimuli and
maximizing contact with others. Contact is minimized when an animal experiences
fear or anxiety, but sometimes an animal avoids a stimulus that is not fear arousing.
Why? And why do animals maintain contact with other stimuli?
A simple answer is that animals maintain contact with stimuli that they find rewarding
in some way and ignore or avoid stimuli that they find neutral or aversive. According
to this view, reward is a mechanism that evolved to help increase the adaptive
fitness both of entire species and of individual members of a species.
But what exactly is reward? One rather circular definition is that reward is the activity
of neural circuits that function to maintain an animal’s contact with certain environmental
stimuli, either in the present or in the future. Presumably, an animal perceives the activity
of these circuits as pleasant. This pleasantness would explain why reward can help
maintain not only adaptive behaviors such as feeding and sexual activity but also potentially
nonadaptive behaviors such as drug addiction. After all, evolution would not have
prepared the brain specifically for the eventual development of psychoactive drugs.
The first clue to the presence of a reward system in the brain came with an accidental
discovery by James Olds and Peter Milner in 1954. They found that rats would perform
behaviors, such as pressing a bar, to administer a brief burst of electrical stimulation
to specific sites in their brains.This phenomenon is called intracranial self-stimulation or
brain-stimulation reward.
Typically, rats will press a lever hundreds or even thousands of times per hour to
obtain this brain stimulation, stopping only when they are exhausted.Why would animals
engage in such a behavior when it has absolutely no value to their survival or to
that of their species? The simplest explanation is that the brain stimulation is activating
the system underlying reward (Wise, 1996).
After 50 years of research on brain-stimulation reward, investigators now
know that dozens of sites in the brain will maintain self-stimulation. Significantly,
however, some regions, including the lateral hypothalamus and medial forebrain
bundle, are especially effective. Stimulation there activates fibers that form the ascending
pathways from dopamine-producing cells of the midbrain tegmentum,
shown in Figure 11-29. This pathway, as you learned in Chapters 5 and 7, is the
mesolimbic dopamine pathway that sends dopamine-containing terminals to various
sites, including especially the nucleus accumbens and the prefrontal cortex.
Neuroscientists believe that the mesolimbic dopamine system is central to
circuits mediating reward for several reasons:
1. Dopamine release shows a marked increase when animals are engaged in intracranial
self-stimulation.
2. Drugs that enhance dopamine release increase self-stimulation,whereas drugs that
decrease dopamine release decrease self-stimulation. It seems that the amount of
dopamine released somehow determines how rewarding an event is.
3. When animals engage in behaviors such as feeding or sexual activity, the release of
dopamine rapidly increases in locations such as the nucleus accumbens.
4. Highly addictive drugs such as nicotine and cocaine increase the level of dopamine
in the nucleus accumbens.
Even opiates appear to effect at least some of an animal’s actions through the dopamine
system.Animals quickly learn to press a bar to obtain an injection of opiates directly into
the midbrain tegmentum or the nucleus accumbens. The same animals do not work to
obtain the opiates if the dopaminergic neurons of the mesolimbic system are inactivated.
Apparently, then, animals engage in behaviors that increase dopamine release.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 433
On the CD, watch a video showing
self-stimulation in a rat in the area on
electrical stimulation in the module on
Research Methods.
Ventral
tegmenum
Caudate nucleus
Nucleus
accumbens in
basal ganglia
Prefrontal
cortex
Cerebellum
Substantia
nigra
Figure 11-29
Mesolimbic Dopamine System Axons
emanating from the ventral tegmentum
(blue arrows) project diffusely through
the brain to synapse on their targets.
Dopamine release in the mesolimbic
pathways has a role in feelings of
reward and pleasure. The nucleus
accumbens is a critical structure in this
“reward system.”
CH11.qxd 2/3/05 4:20 PM Page 433
ls
le
ll
Note, however, that dopamine is not the only reward compound in the brain. For
example, Rainer Spanagel and Friedbert Weiss (1999) stressed that drugs can be rewarding
in the absence of dopamine, and Keith Trujillo and his colleagues (1993)
found that the reinforcing actions of opiates take place through activation of both
dopaminergic and nondopaminergic systems. These findings suggest the existence of
more than one reward-related system in the brain.
In Chapter 7, we encountered the idea that reward has multiple parts in our consideration
of Robinson and Berridge’s incentive-sensitization theory of addiction. These
researchers proposed that reward contains separable psychological components, corresponding
roughly to “wanting,” which is often called incentive, and “liking,” which is
equivalent to an evaluation of pleasure (Robinson & Berridge, 1993, 2003).This idea can
be applied to discovering why we increase contact with a stimulus such as chocolate.
Two independent factors are at work: our desire to have the chocolate (wanting) and
the pleasurable effect of eating the chocolate (liking). This distinction is important. If
we maintain contact with a certain stimulus because dopamine is released, the question
becomes whether the dopamine plays a role in the wanting or the liking aspect of the
behavior. Robinson and Berridge proposed that wanting and liking processes are mediated
by separable neural systems and that dopamine is the transmitter in wanting. Liking,
they hypothesize, entails opioid and benzodiazepine–GABA systems.
According to Robinson and Berridge, wanting and liking are normally two aspects
of the same process; so rewards are usually wanted and liked to the same degree.
However, it is possible, under certain circumstances, for wanting and liking to change
independently. Consider rats with lesions of the ascending dopaminergic pathway
to the forebrain.
These rats do not eat. Is it simply that they do not desire to eat (a loss
of wanting) or has food become aversive to them (a loss of liking for it)? To
find out which factor is at work, the animals’ facial expressions and body
movements in response to food can be observed to see how liking is affected.
After all, when animals are given various foods to taste, they produce
different facial and body reactions, depending on whether they perceive the
food as pleasant or aversive.
For example, when a normal person tastes something sweet, he or she
usually responds by licking the fingers or the lips, as shown at the top of
Figure 11-30. In contrast, if the taste is unpleasantly salty, say, as shown in
the bottom panel, the reaction is often spitting, grimacing, or wiping the
mouth with the back of the hand. Rats, too, show distinctive positive and
negative responses to pleasant and unpleasant tastes.
So, by watching these responses when food is squirted into the mouth
of a rat that otherwise refuses to eat, we can tell to what extent a loss of liking
for food is a factor in the animal’s food rejection. Interestingly, rats that
do not eat after receiving lesions to the dopamine pathway act as though
they still like food.
Now consider a rat with a self-stimulation electrode in the lateral hypothalamus.
This rat will often eat heartily while the stimulation is on. The
obvious inference is that the food must taste good—presumably even better
than it does usually. But what if we squirt food into the rat’s mouth and
observe its behavior when the stimulation is on versus when it is off?
If the brain stimulation primes eating by evoking pleasurable sensations,
we would expect that the animal would be more positive in its facial
and body reactions toward foods when the stimulation is turned on. In
fact, the opposite is found. During stimulation, rats react more aversively
to tastes such as sugar and salt than when stimulation is off. Apparently,
the stimulation increases wanting but not liking.
434 ! CHAPTER 11
Positive reactions
Negative reactions
Figure 11-30
Human Reactions to Taste Positive
(hedonic) reactions are elicited by
sucrose and other palatable tastes.
Hedonic reactions include licking the
fingers and licking the lips. Negative
(aversive) reactions are elicited by
quinine and other nonpalatable tastes.
Aversive reactions include spitting,
making a face of distaste, and wiping
the mouth with the back of the hand.
Adapted from “Food Reward: Brain Substrates of
Wanting and Liking,” by K. C. Berridge, 1996,
Neuroscience and Biobehavioral Reviews, 20, p. 6
CH11.qxd 2/3/05 4:20 PM Page 434
ls
le
ll
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 435
In conclusion, experiments of this sort show that what appears to be a single event—
reward—is actually composed of at least two independent processes. Just as our visual system
independently processes “what” and “how” information in two separate streams, our
reward system appears to include independent processes of wanting and liking.Reward is
not a single phenomenon any more than the processes of perception or memory are.
SUMMARY
How do we experience emotions and motives? Our inner, subjective feelings and
thoughts influence how we behave.We interpret these subjective feelings cognitively as
a range of emotion, as love at one end and hate at the other.Motivation is the cause of
behavior that seems purposeful.As with emotion, we describe motivation in subjective
terms such as hunger and curiosity. Both emotions and motivations are inferred states
that can be without awareness or intent. Unconscious emotions and motives make it
difficult to conclude that our behavior is controlled by the state that we call free will.
What controls the behaviors in which animals will engage at any given time? Animals
are motivated to engage in behavior for its reward. Rewards range from sensory stimulation
to the activity of hormones to the activity of dopamine cells in the brainstem.
Neural circuits controlling species-typical behaviors such as mouse killing by cats are
organized in the brainstem and manifest the evolutionary advantage for these active
circuits: they are rewarding. If animals did not want to engage in motivated behaviors,
their species would become extinct.
How do evolution and experience interact with neurobiology to produce behavior? Behavior
is controlled by its consequences as well as by its biology. Consequences may affect
the evolution of the species or the behavior of an individual animal. Behaviors that
are selected by evolution are often triggered by innate releasing mechanisms. Behaviors
that are selected only in an individual animal are shaped by that animal’s environment
and are learned.
What are the chemical senses and how do they function? Smell and taste are based on
the detection of chemical signals by the olfactory and gustatory senses. Chemical neuroreceptors
interact with chemosignals, leading to neural activity in cranial nerve 1 for
olfaction and cranial nerves 7, 9, and 10 for taste. The cranial nerves enter the brain
and, through a series of synapses, pass to brainstem and forebrain areas. Smell and taste
input merges in the orbitofrontal cortex to produce our perception of flavor.
What principal neural circuit operates in emotional and motivated behavior? The neural
structures that initiate emotional and motivated behaviors are the hypothalamus,
the pituitary gland, the amygdala, the dopamine and noradrenaline pathways from nuclei
in the lower brainstem, and the frontal lobes.
What stimulates emotional and motivated behavior? The experience of both emotion
and motivation is controlled by activity in the ANS, hypothalamus, and forebrain,
In Review .
Rewards are the effects that events have on the behavior of animals. Neural circuits maintain
contact with rewarding environmental stimuli in the present or in the future through
liking and wanting subsystems. The challenge for researchers, as well as their reward, lies
in separating the neural subsystems that take part in reward and in accounting for how the
rewarding effects of environmental events influence liking or wanting.
CH11.qxd 2/3/05 4:20 PM Page 435
ls
le
ll
especially the amygdala and frontal cortex. Emotional and motivated behavior may be
an unconscious response to internal or external stimuli and be controlled by the activity
of innate releasing mechanisms or be a cognitive response to events or thoughts.
What is the difference in the neural control of regulatory and nonregulatory behaviors?
The two distinctly different types of motivated behaviors are (1) regulatory (homeostatic)
behaviors that maintain vital body-system balance and (2) nonregulatory behaviors,
basically consisting of all other behaviors. These nonregulatory behaviors are
not controlled by a homeostatic mechanism and are not reflexive. Feeding is a regulatory
behavior controlled by the interaction of the digestive and hormonal systems and
the hypothalamic and cortical circuits. Sexual activity is a nonregulatory behavior motivated
by the amygdala. Copulatory behavior is controlled by the hypothalamus (the
ventromedial hypothalamus in females and the preoptic area in males).
Why do we find certain behaviors rewarding? Survival depends on maximizing contact
with some environmental stimuli and minimizing contact with others. Reward is
a mechanism for controlling this differential. Two independent features of reward are
wanting and liking. The wanting component is thought to be controlled by dopaminergic
systems, whereas the liking component is thought to be controlled by opiate–
benzodiazepine systems.
KEY TERMS
REVIEW QUESTIONS
1. What are some causes of behavior?
2. Compare the evolutionary and environmental influences on behavior.
3. What key brain structures control motivated behavior? How does each
contribute to this control?
4. Contrast the organizing and activating effects of hormones.
5. Contrast the roles of the hypothalamus and the amygdala in sexual behavior and
sexual motivation.
6. Compare the organization and operation of the chemical senses with those of
vision.
7. Consider the similarities and differences between the brain’s control of
emotional pain and physical pain.
amygdala, p. 411
androgen, p. 393
anorexia nervosa, p. 423
aphagia, p. 424
drive, p. 393
emotion, p. 390
evolutionary psychology,
p. 396
generalized anxiety
disorder, p. 421
hippocampus, p. 411
homeostatic mechanism,
p. 405
hyperphagia, p. 424
hypovolumic thirst, p. 426
innate releasing
mechanism (IRM),
p. 395
Klüver-Bucy syndrome,
p. 416
learned taste aversion,
p. 402
medial forebrain bundle
(MFB), p. 406
motivation, p. 390
nonregulatory behavior,
p. 405
obesity, p. 421
osmotic thirst, p. 426
panic disorder, p. 421
pheromone, p. 399
phobia, p. 421
pituitary gland, p. 406
prefrontal cortex, p. 412
preparedness, p. 402
psychosurgery, p. 418
regulatory behavior, p. 405
reinforcer, p. 400
releasing hormone, p. 408
sensory deprivation, p. 391
sexual dimorphism, p. 428
sexual identity, p. 431
sexual orientation, p. 431
somatic marker hypothesis,
p. 416
436 ! CHAPTER 11
neuroscience interact ive
Many resources are available for
expanding your learning on-line:
www.worthpublishers.com/kolb/
chapter11
Try some self-tests to reinforce your
mastery of the material. Look at some
of the updates on current research on
the brain. You’ll also be able to link to
other sites that will reinforce what
you’ve learned.
www.adaa.org
Link to this Web site to learn more
about anxiety disorders.
www.aabainc.org
Learn more about the research and
treatment of anorexia nervosa and
bulimia at this Web site of the
American Anorexia and Bulimia
Association.
On your CD-ROM, in the module on
the Central Nervous System, you can
review some of the anatomical
structures that are important to
understanding what causes behaviors.
CH11.qxd 2/3/05 4:20 PM Page 436
ls
le
ll
FOR FURTHER THOUGHT
1. Why do cats kill birds? Use the same logic to account for a specific human
behavior of your choice.
2. How could a concept such as preparedness explain racism?
3. What can you infer about brain and behavior relations from the finding that
stimulation of the hypothalamus elicits complex behaviors such as feeding,
digging, and sexual activity?
4. What are the social and moral implications of evidence that sexual orientation is
associated with brain organization?
RECOMMENDED READING
Barondes, S. H. (1993).Molecules and mental illness. New York: Scientific American Library.
A very readable summary of the neurochemical bases of various forms of psychiatric
disease. The Scientific American Library series has excellent illustrations and is written
for an educated lay audience.
Becker, J. B., Breedlove, S. M., & Crews, D. (2002). Behavioral endocrinology (2nd ed.).
Cambridge, MA: MIT Press.What is behavioral endocrinology and why study it? This
book provides the answers. It is a broad survey of the effects of hormones on the
behavior of humans and other animals. The topics range from sexual behavior to
cognitive and motor behaviors.
Damasio, A. R. (1999). The feeling of what happens: Body and emotion in the making of
consciousness. New York: Harcourt Brace. Damasio argues that emotions are curious
adaptations that are part and parcel of the neural machinery that we have evolved for
our survival. Damasio’s ideas have developed from his study of people with frontal-lobe
injuries who have abnormal emotional control and a parallel abnormality in other
cognitive activities. This interesting book has influenced thinking about the role of
emotion in the brain’s daily activities.
Eibl-Eibesfeldt, I. (1989).Human ethology. New York: Aldine de Gruyter. One of the first
ethologists to study human behavior, Eibl-Eibesfeldt has written a thorough book in
which he summarizes what is known about the species-typical behavior of people. This
book provides a wealth of photographic examples of human behaviors that are
genetically programmed and found throughout the world’s cultures.
Lane, R. D., & Nadel, L. (Eds.). (2000). Cognitive neuroscience of emotion. New York: Oxford
University Press. This book is a showcase for the newly emerging ideas of the cognitive
neuroscience of emotion. The chapters range from heavily theoretical accounts to
strongly empirical ones, but all focus on the role of emotion in cognition.
Robinson, T. E., & Berridge, K. C. (1993). The neural basis of drug craving: An incentivesensitization
theory of addiction. Brain Research Reviews, 18, 247–291. The Robinson-
Berridge theory of drug addiction and craving is a thorough analysis of the evidence
that wanting and liking drugs are two different things that likely have different neural
bases.
Woods, S. C., Seeley, R. J., Porte, D., & Schwartz, M.W. (1998). Signals that regulate food
intake and energy homeostasis. Science, 280, 1378–1382. Obesity is an increasingly
prevalent and important health problem. Naturally, those who wish to lose weight are
hoping that a magic bullet will be found to treat obesity. This article is one in a special
issue of Science that looks at the regulation of body weight. The authors review the
signals that tell us when to eat (or not) and conclude that a single magic bullet is
unlikely but that treatments aimed at multiple targets may be realistic.
WHAT CAUSES EMOTIONAL AND MOTIVATED BEHAVIOR? ! 437
CH11.qxd 2/3/05 4:20 PM Page 437
Focus on Comparative Biology: The Variety of
Biological Rhythmns
A Clock for All Seasons
Origins of Biological Rhythms
Biological Clocks
Biological Rhythms
Free-Running Rhythms
Zeitgebers
Focus on Disorders: Seasonal Affective Disorder
Neural Basis of the Biological Clock
Suprachiasmatic Rhythms
Evidence for Dual Clocks
Immortal Time
What Ticks?
Pacemaking Circadian Rhythms
Focus on New Research: Synchronizing Biorhythms at
the Molecular Level
Pacemaking Circannual Rhythms
Sleep Stages and Dreaming
Measuring How Long We Sleep
Measuring Sleep in the Laboratory
A Typical Night’s Sleep
Non-REM Sleep
Focus on Disorders: Restless Legs Syndrome
REM Sleep and Dreaming
What We Dream About
What Does Sleep Accomplish?
Sleep As a Passive Process
Sleep As a Biological Adaptation
Sleep As a Restorative Process
Sleep and Memory Storage
Neural Basis of Sleep
Reticular Activating System and Sleep
Neural Basis of the EEG Changes Associated
with Waking
Neural Basis of REM Sleep
Sleep Disorders
Disorders of Non-REM Sleep
Focus on Disorders: Sleep Apnea
Disorders of REM Sleep
Sleep and Consciousness
438 !
C H A P T E R 12
Why Do We Sleep and Dream?
Left: Bob Thomas/Tony Stone. Middle: Dr. Dennis Kunkel/Phototake.
Right: Hank Morgan/Photo Researchers.
ls
le
ll
CH12.qxd 2/7/05 2:58 PM Page 438
Sea-bear behavior is remarkable to us humans. We are
diurnal animals (from the Latin dies, meaning “day”): we are
active during daylight, and we sleep when it is dark. Our recent
evolutionary history places early modern humans in
Africa at equatorial latitudes, where day and night are almost
equal in length. Because we are adapted for daylight
vision and have difficulty seeing well at night, we humans
are not adapted by natural selection for darkness. That animals
seek out darkness and flourish in it seems strange to us.
We sleep for about 8 hours each night on average. Our
sleep is characterized by a decline in body temperature and
a loss of awareness of our surroundings. Sea bears sleep in
the winter, as mentioned, but their summer torpor seems related
to hibernation. The strategy used by rodents to extend
body-fat supplies as long as possible is similar to shallow
torpor except that declines in body temperature are so extreme
that the animals expend almost no energy.
Despite the great differences among human, bear, and
rodent behaviors, all are adaptations that maximize obtaining
food and to minimize spending or losing energy stores
obtained from food. Humans evolved to be active during
the day—we see best then to obtain food—and inactive at
night—to conserve body resources. Sea bears hunt all winter
and so build up fat supplies. They enter torpor and so
extend the period during which they live on those fat stores
when resources are scarce.
Sea bears are clearly prepared to go out on the ice well
in advance of its formation. They walk along the Arctic
shores for weeks before the ice forms; they also leave
the ice before it melts. Birds migrate before food resources
are depleted and winter arrives. Rodents gorge themselves,
build nests, and store food in their burrows before winter
and hibernation set in. The migratory behavior of birds also
is a strategy used to maximize food acquisition and minimize
energy loss, except the objective is achieved by moving
to a habitat where food is abundant.
The Variety of Biological Rhythmns
Focus on Comparative Biology
A s winter approaches in Northern latitudes, many
Arctic animals prepare their escape. Arctic terns fly
15,000 kilometers to Antarctica, where it is summer. Lemmings,
mice, and ground squirrels cannot travel long distances;
these rodents spend the winter in burrows in a
sleeplike state called hibernation.
Polar bears, in contrast, congregate to go out onto the
pack ice. They migrate toward the Arctic as the days grow
ever shorter. Some travel thousands of kilometers. In the
continuous darkness of the Arctic winter, polar bears (also
known as sea bears, Ursus maritimus) hunt seals, walrus,
and whales. They take time to sleep on the ice but, because
their world is continuously dark, their sleep cannot be
called either nighttime or daytime sleep.
When summer comes again, the birds return and the
rodents emerge from their burrows. The sea bears return
from the ice, dig beds in the earth, and spend the entire
summer in shallow torpor—a condition resembling sleep
except that the decline in body temperature is greater than
during sleep. If the bears have access to food throughout
the year, they do not enter torpor. Even if you find torpor
somehow appealing, it remains hard to imagine spending
all summer unconscious.
ls
le
ll
David Myers/Tony Stone
Torpor. Inactive condition resembling
sleep but with a greater decline in body
temperature.
Diurnal animal. Organism that is active
chiefly during daylight.
CH12.qxd 2/7/05 2:58 PM Page 439
ls
le
ll
440 ! CHAPTER 12
We are similar to sea bears, rodents, and birds in one other way. Their behaviors
and ours evolved to respond not simply to immediate environmental
changes but also to longer-term changes that result in food abundance or
shortages. Today, with humans controlling more of their environment, food abundance
or shortage may be a matter of the distance to the nearest pizzaria.
We live by the clock, too, retiring to sleeping sites in preparation for sleep and frequently
arising before it is fully light to prepare for our daily activities. But we, along
with other animals, still appear to have warning of impending winter or impending
changes in the day–night cycle. There are environmental signals to which we all respond.
These signals prime us to anticipate daily and seasonal changes. But, even in the
absence of obvious signals, we and other animals can anticipate daily and seasonal
changes. It follows that we have a biological clock that regulates our behavior in concert
with the rhythmic changes of our world.
In this chapter,we seek answers to questions related to biological rhythms and sleep:
How is our behavior modified to cope with the day–night cycle?
Why has sleep evolved as a strategy for waiting out the night?
What neural mechanisms regulate sleep and waking?
How do disorders of the natural rhythms of sleep occur?
A CLOCK FOR ALL SEASONS
We first consider evidence for the existence of a biological clock, how that clock keeps
time, and how it regulates our behavior. Because environmental cues themselves are
not always consistent, we examine the role of biological clocks in interpreting environmental
cues in an intelligent way.
Origins of Biological Rhythms
Biorhythms, the inherent rhythms that control or initiate various biological processes,
are linked to the rhythmic cycle of days and seasons produced by Earth’s rotation on
its axis and by its progression in orbit around the sun (Figure 12-1). Earth rotates on
Biorhythm. Inherent rhythm that
controls or initiates various biological
processes.
Seasons change as the
earth revolves around
the sun. Each revolution
takes 1 year.
Day and night result from rotation of
the earth on its axis every 24 hours.
The tilt of the earth on its
axis determines seasons.
Sun
Northern winter Northern summer
Figure 12-1
Origins of Biorhythms
Each part of the earth faces
the sun for part of its daily
rotation cycle (daytime)
and faces away from the
sun for the other part
(nighttime). Seasonal
changes in temperature
and in the amount of
daylight are related to the
annual revolution of the
earth around the sun and
the tilt of its axis.
CH12.qxd 2/7/05 2:58 PM Page 440
ls
le
ll
its axis once every 24 hours, producing a 24-hour cycle of day and night. Earth’s axis is
tilted slightly, and so, as Earth orbits the sun once each year, the North and South Poles
incline slightly toward the sun for part of the year and slightly away from the sun for
the rest of the year.
When inclined toward the sun, the Southern Hemisphere experiences
summer and gets more direct sunshine for more hours each
day, and the climate is warmer. At the same time, the Northern
Hemisphere, inclined away from the sun, receives less direct sunlight,
making the days shorter and the climate colder. Tropical regions,
being near the equator, undergo little seasonal change as Earth progresses
around the sun.
The seasonal differences in polar and equatorial regions cause
animals living near the poles to be relatively more affected by seasonal
changes and animals living near the equator to be relatively
more affected by day-and-night changes. Seasonal and daily changes
may have combined effects on organisms as well, inasmuch as the
onset and duration of daily changes depend on the season and latitude.
Animals living in polar regions also have to cope with greater
fluctuations in daily temperature, light, and food availability than do
animals living near the equator.
We humans are equatorial animals in that our behavior is governed
more by daily cycles than by seasonal cycles. Our behavior is
dominated by a rhythm of daylight activity and nocturnal sleep. Not
only does human waking and sleep behavior cycle daily, so also do
pulse rate, blood pressure, body temperature, rate of cell division,
blood-cell count, alertness, urine composition, metabolic rate, sexual
drive, and responsiveness to medications.
Biorhythms are not unique to animals. Plants display rhythmic
behavior, exemplified by species in which leaves or flowers open
during the day and close at night. Even unicellular algae and fungi
display rhythmic behaviors related to the passage of the day. Some
animals, including lizards and crabs, change color in a rhythmic
pattern. The Florida chameleon, for example, turns green at night,
whereas its coloration matches its environment during the day. In
short, almost every organism and every cell display changes of some
sort that are related to daily or seasonal changes.
Biological Clocks
If the behavior of animals were affected only by seasonal and daily
changes, the neural mechanisms that account for changes in behavior
would be much simpler to study than they are. Behavior would
be driven by external cues, which would be easy to identify, and, accordingly,
the neural processes that respond to those cues also would
be easy to identify.
That behavior is not driven simply by external cues was first recognized
in 1729 by French geologist Jean Jacques D’Ortous de Mairan
(see Raven, Evert, & Eichorn, 1992). In an experiment similar to that
illustrated in the Procedure section of Experiment 12-1, de Mairan
isolated a plant from daily light and dark cues and from temperature
cues and noted that the rhythmic movements of its leaves continued,
WHY DO WE SLEEP AND DREAM? ! 441
Revolving drum
Pen
1 2 3 4
Days in continuous dim light
Leaf up
Leaf down
Movement of the plant is endogenous. It
is caused by an internal clock that matches
the temporal passage of a real day.
Conclusion
Question: Is plant movement exogenous or endogenous?
EXPERIMENT 12-1
A pen attached to a
leaf is moved when
the leaf moves,…
The movements of the
plant’s leaves are recorded
in constant dim light.
…producing a record
of the movement.
Procedure
Results
Leaf up
Leaf down
Jack Dermid Jack Dermid
CH12.qxd 2/7/05 2:58 PM Page 441
ls
le
ll
as graphed in the Results section of the experiment. What concerned de Marain’s followers
was the possibility that some undetected external cue stimulated the rhythmic
behavior of the plant.
Such cues could include changes in gravity, changes in electromagnetic fields, and
even changes in the intensity of rays from outer space. Nevertheless, further experiments
showed that the daily fluctuations are endogenous—they come from within the
plant. In fact, experiments show that most organisms have an internal biological clock
that matches the temporal passage of a real day.
Your clock and calendar enable you to plan and schedule your time. Your biological
clock performs these functions, too. An endogenous biological clock allows an animal,
in effect, to anticipate events: to migrate before it gets cold rather than waiting
until it gets cold and to mate at the correct time of the year. The clock allows animals
to arrive at the same place at the same time if they are to mate or to begin a migration.
Most important, a biological clock signals to an animal that, if daylight lasts for about
12 hours today, it will last for about 12 hours tomorrow but, if it lasts for 4 hours today,
it will last for about 4 hours tomorrow.
Plants and animals evolved internal clocks through natural selection to avoid being
tricked into displaying maladaptive behavior. Plant bulbs that begin to grow during a
January thaw only to be killed by a subsequent cold spell exemplify such maladaptive
behavior.
Biological Rhythms
Although the existence of endogenous biological clocks was demonstrated more than
200 years ago, the detailed study of biorhythms had to await the development of procedures
that could analyze ongoing behavior over a long period of time. Behavior
analysis requires a method for counting behavioral events and a method for displaying
those events in a meaningful way. For example, the behavior of a rodent can be measured
by giving the animal access to a running wheel, such as that illustrated in Figure
12-2A, in which it can exercise.
A chart recorder or a computer records each turn of the wheel and displays the result
(Figure 12-2B). Because most rodents are nocturnal, sleeping during light hours and becoming
active during dark hours, their wheel-running activity takes place in the dark
(Figure 12-2C). If each day’s activity is plotted under the preceding day’s activity in a column,
we can observe a pattern of activity over a period of time. Various details of the
chart can then be examined, including when the animal was active and how active it was.
442 ! CHAPTER 12
Biological clock. Neural system that
times behavior.
Period. Time required to complete a
cycle of activity.
Animal’s activity
12 noon 6 PM Dark 6 AM 12 noon
(A) (B)
Rat has access to
a running wheel.
Turns of the wheel are
recorded on a chart recorder,
which plots each wheel
rotation as a tick on a chart.
(C)
Each line represents one day’s activity. When
activity was plotted for a month under
conditions of no light between 6:00 PM and
6:00 AM, the rat was shown to be active
during dark hours of the day–night cycle.
Figure 12-2
Recording the Daily Activity Cycle of
a Rat Adapted from Biological Clocks in
Medicine and Psychiatry (pp. 12–15), by C. P.
Richter, 1965, Springfield, IL: Charles C Thomas.
CH12.qxd 2/7/05 2:58 PM Page 442
ls
le
ll
Perhaps the most important piece of information charted by an activity record is
the cycle of activity. The time required to complete a cycle of activity is called a period.
The activity period of most rodents is about 24 hours in an environment in which the
lights go on and off with regularity.Our own sleep–wake period also is about 24 hours.
Many other kinds of behaviors, however, have periods that are more or less than
24 hours.
Animals have a surprising number of biological clocks, and those clocks have varying
periods. Two kinds of rhythms typical of most animals are circannual rhythms
(Latin circa, meaning “about,” plus annuum, meaning “year”), of which the migratory
cycles of sea bears and Arctic terns are examples, and circadian rhythms (from Latin
for “about a day”), which are the day–night rhythms found in almost all animals and
cellular processes. These two are not the only kinds of rhythms, however.
Infradian (Latin infra, meaning “smaller than”) rhythms are those that have a period
of less than one day. Our eating behavior, which takes place about every 90 minutes
to 2 hours, including snacks, is an example of an infradian rhythm. Rodents,
although active throughout the night, are most active at the beginning and end of the
dark period.
Many sea-dwelling animals’ rhythms are about 12 hours,
which matches the twice-daily changes in tides produced by the
pull of the moon on the earth and its oceans. Therefore, an infradian
rhythm is embedded within their circadian rhythm.
Our eye-blink rate, our heart rate, and even the rhythmic action
potentials of some of our neurons are other examples of infradian
rhythms.
Other biorhythms have periods greater than a day and less than a year. These are
ultradian (meaning “beyond a day”) rhythms. The menstrual cycle of female humans,
with an average period of about 28 days, is an ultradian biorhythm.
We focus in this chapter on the circadian rhythm, which is central to our sleep–
waking behavior. Note, however, that the fact that a behavior appears to be rhythmic
does not mean that it is ruled by a biological clock. There is evidence that sea bears will
remain on the ice as long as the ice pack and food supplies last, and many migrating
birds will postpone their migrations as long as they have a food supply. Therefore,
whether a rhythmic behavior is produced by a biological clock must be demonstrated
experimentally. A definitive experiment to support the conclusion that the sea bear
does have a clock would be methodologically difficult to conduct, but such demonstrations
are not difficult to make with other animals, including ourselves.
Free-Running Rhythms
To determine if a rhythm is produced by a biological clock, researchers must design a
test in which they remove all external cues. If light is proposed to be a major cue, the
experiment can be set up in three ways: a test can be given in continuous light, it can
be given in continuous darkness, or the selection of light or darkness can be left to the
subject.
That the human sleep–waking rhythm is governed by a biological clock was first
demonstrated by Jurgen Aschoff and Rutger Weber (see Kleitman, 1965), who allowed
subjects to select their light–dark cycle. The experimenters placed individual subjects
in an underground bunker where no cues signaled when day began or ended. Thus, the
subjects selected the periods when their lights were on or off, when they were active,
and when they slept. In short, they selected their own day and night length. By measuring
ongoing behavior and recording sleeping periods with sensors on the beds,
WHY DO WE SLEEP AND DREAM? ! 443
Circadian rhythm. Day–night rhythm.
Biological rhythm Time frame Example
Circannual Yearly Migratory cycles of birds
Infradian Less than a year Human menstrual cycle
Circadian Daily Human sleep cycle
Ultradian Less than a day Human eating cycles
CH12.qxd 2/7/05 2:58 PM Page 443
ls
le
ll
444 ! CHAPTER 12
Aschoff and Weber found that the subjects continued to show daily sleep-activity
rhythms. This finding demonstrated that humans have an endogenous biological clock
that governs sleep–waking behavior.
Figure 12-3 shows, however, that the biorhythms recorded by Aschoff and Weber
were peculiar when compared with the rhythms before and after isolation. Although
the period of the subjects’ sleep–wake cycles approximated a normal period of 24 hours
before and after the test, during the test they progressively deviated away from clock
time. Rather than being 24 hours, the activity period in the bunker was about 25 to 27
hours, depending on the subject.
The subjects were choosing to go to bed from 1 to 2 hours later every “night.” A
shift by an hour or so of sleeping time is not remarkable for a few days, but its cumulative
effect quickly became dramatic: soon the subjects were getting up at about the
time the experimenters outside the bunker were going to bed. Clearly, the subjects were
displaying their own personal cycles. A rhythm that runs at a frequency of the body’s
own devising when environmental cues are absent is called a free-running rhythm.
The period of free-running rhythms depends on the way in which external cues
are removed.When hamsters, a nocturnal species, are tested in constant darkness, their
free-running periods are a little shorter than 24 hours; when they are tested in constant
light, their free-running periods are a little longer than 24 hours. This test dependency
in hamsters is typical of nocturnal animals.
As Figure 12-4 shows, the opposite free-running periods are typical of diurnal animals
(Binkley, 1990).When sparrows, which are diurnal birds, are tested in constant
darkness, their free-running periods are a little longer than 24 hours; when they are
tested in constant light, their free-running periods are a little shorter than 24 hours.
Why periods change in different lighting conditions is not clear, but a rule of thumb is
that animals expand and contract their sleep periods as the sleep-related lighting period
expands or contracts.
Understanding this point enables you to predict how artificial lighting influences
human circadian periods, and you can offer an explanation of why Aschoff and Weber’s
subjects displayed periods that were longer than 24 hours. Endogenous rhythmicity is
not the only factor that contributes to circadian periods, however. An endogenous
In
bunker
Before
bunker
After
bunker
Hour of day
25
20
15
10
5
1
Dark
period
Light
period
Days
0 8 16 24 8 16 24 8 16 24 8
Waking periods of subjects placed in the
bunker began to change because there
were no cues to light and dark periods. The
purple lines represent daily activity rhythm.
Over a period of time,
subjects in the bunker were
getting up at about the time
experimenters outside the
bunker were going to bed.
Figure 12-3
Free-Running Rhythm in a Human
Subject The record for days 1 through
3 shows the daily sleep period under
normal day–night conditions. That for
days 4 through 20 shows the freerunning
rhythm that developed while
the subject was isolated in a bunker and
allowed to control day and night length.
The daily activity period shifts from 24
hours to 25.9 hours. On days 21 through
25, the period returns to 24 hours when
the subject is again exposed to a normal
light-and-dark cycle. Adapted from Sleep
(p. 33), by J. A. Hobson, 1989, New York: Scientific
American Library.
Free-running rhythm. Rhythm of the
body’s own devising in the absence of all
external cues.
CH12.qxd 2/7/05 2:58 PM Page 444
ls
le
ll
WHY DO WE SLEEP AND DREAM? ! 445
rhythm that is just a little slow or a little fast would be useless because the error would
accumulate, and so there must also be a mechanism for setting rhythms to correspond
to environmental events.
Zeitgebers
Because Aschoff and Weber’s subjects had a sleep–wake cycle of 24 hours before and
after they entered the experiment and because hamsters usually have a 24-hour rhythm,
we might wonder how normal rhythms are maintained. The biological clock must keep
to a time that matches changes in the day–night cycle. If a biological clock is like a
slightly defective wristwatch that runs either too slow or too fast, it will eventually provide
times that are inaccurate by hours and so become useless.
If we reset the wristwatch each day, however—say, when we awaken—it would
then provide useful information even though it is not perfectly accurate. There must
be an equivalent way of resetting a free-running biological clock. In experiments to determine
how clocks are set, researchers have found that cues such as sunrise and sunset,
eating times, and other activities can all set the circadian clock.
Normally, light is the most potent stimulus. Aschoff and Weber called such cues
Zeitgebers (German for “time givers”).When a clock is reset by a Zeitgeber, it is said to
be entrained. The importance of light in entraining circadian rhythms is explained in
“Seasonal Affective Disorder.”
Biological clocks are flexible.They can be reset each day so that they accurately correspond
to the season. In polar regions, you’ll recall, the time of onset and the length of day
and night are changing as the seasons progress. At the higher latitudes, daylight begins
very early in the morning in summer and very late in the morning in winter. To adjust
to these changes, an animal responds both to daylight and to how long the day will last.
House sparrow
Constant darkness for 18 days
Constant light for 16 days
When tested in constant
darkness, sparrows (which
are diurnal birds) extend
their free-running periods
to a little more than 24
hours.
Each line
represents a day.
When tested in constant
light, sparrows contract
their free-running
periods to a little less
than 24 hours and are
much more active
throughout the
testing period.
Blips represent
hops on a perch.
Figure 12-4
Free-Running Rhythms of a Diurnal
Animal Adapted from The Clockwork
Sparrow (p. 16), by S. Binkley, 1990, Englewood
Cliffs, NJ: Prentice Hall.
Zeitgeber. Environmental event that
entrains biological rhythms; a “time
giver.”
Entrainment. Determination or
modification of the period of a biorhythm.
CH12.qxd 2/7/05 2:58 PM Page 445
ls
le
ll
A biological clock that is reset each day tells an animal that daylight will begin tomorrow
at approximately the same time that it began today and that tomorrow will last
approximately as long as today did. But when should the clock be reset? The current research
opinion is that light Zeitgebers work best when exposure occurs near the beginning
or the end of the light segment of the cycle.
The very potent entraining effect of light Zeitgebers is illustrated by laboratory
studies of Syrian hamsters, perhaps one of the most compulsive animals with respect
to timekeeping.When given access to running wheels, these hamsters exercise during
the night segment of the laboratory day–night cycle. A single brief flash of light is an
effective Zeitgeber for entraining their biological clocks. (If a hamster happens to blink
during this Zeitgeber, the light will still penetrate its closed eyelids and entrain its biological
clock.)
446 ! CHAPTER 12
Seasonal Affective Disorder
Focus on Disorders
In diurnal species, the perception of longer nights by the circadian
pacemaker most likely stimulates pressure for more
sleep. If not satisfied, cumulative sleep deprivation can result.
In seasonal affective disorder (SAD), the light phase of
the circadian rhythm is too short to entrain the circadian
rhythms. Consequently, a person’s biorhythm probably becomes
a free-running rhythm.
People vary in the duration of their free-running rhythm
phases relative to the actual day–night cycle. Some are phase
retarded, with sleep time coming earlier each day; some are
phase delayed, with sleep time coming later each day. Common
symptoms are observed in or reported by more than
two-thirds of people who report that they are depressed during
the winter months in northern latitudes, including
Sadness Later waking
Anxiety Increased sleep time
Irritability Interrupted, not
Decreased physical refreshing
sleep activity Daytime drowsiness
Increased appetite Decreased libido
Carbohydrate craving Menstrual difficulties
Increased weight Work difficulties
Earlier sleep onset Interpersonal difficulties
Researchers consistently report that light is capable of
ameliorating the depression of SA D, and one treatment for
SA D symptoms, phototherapy, uses light to entrain the circadian
rhythm. The basic idea is to increase the short winter
photoperiod by exposing a person to artificial bright
light. Typical room lighting is not sufficiently bright.
A person undergoing phototherapy for SA D sits in front
of a bank of bright lights. Some manufacturers recommend
exposure in the morning, others in the evening. Still others
recommend bracketing the day with morning and evening
exposure. There may be merit to all these recommendations.
Recent evidence suggests that there may be two SC N
oscillators, one regulating activity and one regulating other
body rhythms, each of which may be related to different
Zeitgebers.
Hamster
Exposure to a bright light each winter morning and/or evening
entrains the circadian rhythm. Courtesy of Bio-Light by Enviro-Med, 800-
222-3296, www.bio-light.com
CH12.qxd 2/7/05 2:58 PM Page 446
ls
le
ll
Considering the somewhat less compulsive behavior that most of us display, we
should shudder at the way in which we entrain our own clocks when we stay up late in
artificial light, sleep late some days, and get up early by using an alarm clock on other
days. Such inconsistent behavior with respect to the human biological clock has been
associated with accidents and daytime fatigue.
Entrainment also works best if the adjustment to the biological clock is not too
large. People who work shifts are often subject to huge adjustments, especially when
they work the graveyard shift (11:00 PM to 7:00 AM), the period when they would normally
sleep. The results of studies show that adapting to such a change is difficult and
stressful.Adaptations to shift work are better if people first work the evening shift (3:00
PM to 11:00 PM) for a time before beginning the graveyard shift.
Long-distance air travel—say, from North America to Europe or Asia—also demands
a large and difficult time adjustment. For example, travelers flying east from
New York to Paris will begin their first European day just when their biological clocks
are prepared for sleep (Figure 12-5). The difference between a person’s circadian
rhythm and the daylight cycle in a new environment can produce the feeling of disorientation
called jet lag. The west-to-east traveler generally has a more difficult adjustment
than does the east-to-west traveler, who needs to stay up only a little longer
than normal.
WHY DO WE SLEEP AND DREAM? ! 447
West-to-east travel
(6-hour flight)
Traveler leaves New York at
9:00 PM, when it’s 3:00 AM in
Paris.
1
Traveler arrives in Paris at 9:00 AM, when it’s 3:00 AM in New York.
Because his biological clock is set for New York time, he is prepared
for sleep, whereas residents of Paris are waking.
2
New York (9 PM)
New York (3:00 AM) Paris (9:00 AM)
East-to-west travel
(6-hour flight)
Paris (9:00 PM) New York (3:00 PM)
Traveler leaves Paris at 9:00 PM, when
it’s 3:00 PM in New York.
3
Traveler arrives in New York at 9:00 PM and
needs to stay up only a little longer than
normal for his biological clock to adjust.
4
Paris (3:00 AM)
New York (9:00 PM)
New York (9:00 PM)
Paris (3:00 AM)
Figure 12-5
Jet Lag Disruption in the entrainment
of a person’s biological clock that may
be brought on by jet travel is likely more
pronounced in west-to-east travel,
because the disruption in the person’s
circadian rhythm is dramatic. On the
return journey, the traveler’s biological
clock has a much easier adjustment to
make.
CH12.qxd 2/7/05 2:58 PM Page 447
ls
le
ll
NEURAL BASIS OF THE BIOLOGICAL CLOCK
Curt Richter (1965) was the first researcher to attempt to locate biological clocks in the
brain. In the 1930s, he captured wild rats and tested them in activity wheels. He found
that the animals ran, ate, and drank when the lights were off and were relatively quiescent
when the lights were on. Richter’s hypothesis was that the rats’ behavior and the
biological clock that was responsible for rhythmicity were separate.
Richter proposed that the biological clock acted as a pacemaker to instruct other
neural structures when they should produce the behaviors for which they were responsible.
Thus, behaviors such as running, eating, drinking, and changes in body temperature
occur when the pacemaker tells their relevant neural areas that it is time to begin.
Richter further proposed that this biological clock is localized in the brain, rather
than being a property of all body or all brain cells.By inserting an electrode into the brain
to damage brain tissue with electric current, he found that animals lost their circadian
rhythms after damage to the hypothalamus. Subsequently, by making much more
discrete lesions, experimenters have shown that a region called the suprachiasmatic
nucleus (SCN) acts as a biological clock (Ralph & Lehman, 1991). As illustrated in Figure
12-6, the SCN is located in the hypothalamus, just above (supra) the optic chiasm—
hence its name.
The SCN receives information about light through its own special visual pathway,
the retinohypothalamic pathway. This pathway consists of a subset of cone receptors
in the retina that are connected to a subset of optic-tract fibers and use glutamate as
their primary neurotransmitter in the SCN. Light signals are carried by this pathway
to the suprachiasmatic nucleus to excite and to entrain it. Visual fibers carrying information
about light also go to an area of the thalamus (the intergeniculate leaflet), but
we will limit our consideration of rhythms to the role of the SCN.
Scientists have also found pacemakers in the retina and in the pineal gland. Some
behaviors may be paced by widely distributed pacemaker brain cells. Among the other
possible pacemakers, the pineal gland has received the most study. It acts as a pacemaker
in some species of birds. It is excited by light that enters the brain not through
the visual system but through the skull.When the heads of such birds are painted black,
the pineal gland’s pacemaker activities are blocked. Because the pineal gland can respond
directly to light, it has been called the “third eye.” In most animals, however, the
suprachiasmatic nucleus is the main pacemaker.
Suprachiasmatic Rhythms
Further evidence for the role of the SCN in circadian rhythms comes from a remarkable
series of experiments demonstrating that the neurons of the nucleus have intrinsic
rhythmic activity (Earnest et al., 1999). Following up on Richter’s original experiments,
In Review .
Many behaviors occur in a rhythmic pattern in relation to time. These biorhythms may display
a yearly cycle (circannual rhythms) or a daily cycle (circadian rhythms). Biological
rhythms are timed internally by regions of the nervous system that serve as biological clocks
to regulate most of our circadian rhythms, especially our sleep–wake cycles. Although biological
clocks keep fairly good time, their periods may be slightly shorter or longer than
a 24-hour day unless they are reset each day. Their spontaneous periods are called freerunning
rhythms. Zeitgebers are environmental cues that reset the biological clock.
448 ! CHAPTER 12
Optic
chiasm
Suprachiasmatic
nucleus
Hypothalamus
Figure 12-6
Suprachiasmatic Nucleus in a Rat Brain
Suprachiasmatic nucleus (SCN).
Main pacemaker of circadian rhythms
located just above the optic chiasm.
Retinohypothalamic pathway.
Neural route from a subset of cone
receptors in the retina to the
suprachiasmatic nucleus of the
hypothalamus; allows light to entrain
the rhythmic activity of the SCN.
CH12.qxd 2/7/05 2:58 PM Page 448
ls
le
ll
investigators have found that, if the suprachiasmatic nuclei are selectively lesioned in rodents,
the animals still eat, drink, exercise, and sleep a normal amount, but at haphazard
times.
By itself, disorganized behavior does not definitively demonstrate that the SCN is
the clock that gives instructions about when these activities should take place. The SCN
could just be a way station between receptors in the eye and a clock located elsewhere
in the brain. Three other lines of evidence do show, however, that the suprachiasmatic
nucleus is indeed a biological clock:
1. The metabolic activity of the SCN is higher during the light period of the day–
night cycle than it is during the dark period of the cycle. If 2-deoxyglucose—a
form of glucose that is taken up by metabolically active cells but is not used by
them and cannot escape from them—is tagged with a radioactive label, cells that
are more active will subsequently give off more radioactivity. When 2-deoxyglucose
is injected into rodents, its accumulation by the SCN should be relative to the
animal’s daily rhythm if the neurons in the nucleus are responsible for the rhythm.
More tracer is found in the SCN after injections given in the light period of the
light–dark cycle than after injections given in the dark period. This experiment
demonstrates that suprachiasmatic cells have rhythmic metabolic activity and that
the SCN is special in this respect.
2. Recording electrodes placed in the SCN confirm that neurons in this region are
more active during the light period of the cycle than during the dark period.
3. If all the pathways into and out of the SCN are cut, the neurons of the suprachiasmatic
nucleus maintain their rhythmic electrical activity.
Together, the results of these experiments show that the suprachiasmatic neurons have
a rhythmic pattern of activity that is intrinsic.
Evidence for Dual Clocks
After scientists had demonstrated that the SCN is rhythmically active, the question of
how that rhythmicity is generated became central.When the SCN was removed from
the brain, maintained in a laboratory dish, and subjected to electrical recording, the
neurons were found to maintain their rhythmic activity. Furthermore, if the neurons
were isolated from one another, each one was rhythmic. Individual cells did seem to
have slightly different rhythms, however. This cellular individuality suggests either that,
collectively, the cells express average rhythm or that the SCN has components that are
able to produce rhythms with different periods or both. For example, findings from
studies on the genes that control rhythms in fruit flies suggest that two separate groups
of circadian neurons may control the rhythm of a fly’s evening and morning locomotor
peaks (Stoleru et al., 2004).
Evidence for two oscillators within the SCN have been obtained in the rat (de la
Iglesia et al., 2004). One of the oscillators receives direct projections from the retina,
whereas the other does not. The presence of dual oscillators might explain why all of the
body’s rhythms do not change together in response to temporal changes in Zeitgebers.
For example, rat activity levels change rapidly in response to changes in the onset
of light, whereas change in body temperature rhythm is slower. Applied to humans,
this finding might mean that sleep cycles adapt rapidly for a traveler arriving in
Europe from North America, whereas the body-temperature cycle that accompanies
the sleep–wake period lags behind. The traveler might get a good night’s sleep but feel
tired because his body temperature is low and remains set to accompany pretravel sleep
time.
WHY DO WE SLEEP AND DREAM? ! 449
CH12.qxd 2/7/05 2:58 PM Page 449
ls
le
ll
Immortal Time
How do suprachiasmatic cells develop their rhythmic activity? One possibility is that
the endogenous rhythm is learned. That is, the cells may initially have no rhythm but,
after they receive their first exposure to rhythmic stimulation from environmental Zeitgebers,
they become rhythmic.
One way of examining whether rhythmicity is learned is to maintain animals from
birth in an environment without Zeitgebers. In experiments in which animals are
raised in constant darkness, their behavior still becomes rhythmic. It is possible that
the animals’ fetal suprachiasmatic cells acquired rhythmicity from the mother, but, in
experiments in which animals have been maintained without entraining
cues for a number of generations, each generation continues to display
rhythmic behavior. Even if the mother has received a lesion of the SCN so
that her behavior is not rhythmic, the behavior of the offspring is rhythmic.
Thus, it seems that rhythmicity is not learned.
A line of evidence supporting the idea that suprachiasmatic cells are
genetically programmed for rhythmicity comes from studies performed in
Canada by Martin Ralph and his coworkers with the use of transplantation
techniques (Ralph & Lehman, 1991). The general design of the experiments
is illustrated in Figure 12-7. First, hamsters are tested in constant
dim light or in constant darkness to establish their free-running rhythm.
They then receive a suprachiasmatic lesion, followed by another test to
show that the lesion has abolished their rhythmicity. Finally, the hamsters
receive transplants of suprachiasmatic cells obtained from hamster embryos.
About 60 days later, the hamsters again show rhythmic activity,
demonstrating that the transplanted cells have become integrated into the
host brain and are responsible for reestablishing rhythmic behavior.
In further experiments, Ralph and his coworkers identified and selectively
bred hamsters that displayed a 20-hour rhythm. They named the
gene that was responsible for the short rhythm tau. If they destroyed the
SCN in a genetically normal hamster with a 24-hour period and then
transplanted cells from a fetal 20-hour hamster into the cavity, the former
24-hour hamster exhibited the 20-hour period of the hamster having the
tau gene.
David Earnest and his coworkers (1999) carried the transplantation
methodology one step further. They harvested cells from the rat SCN and
used them to produce an immortalized cell line. By treating each generation
of cells with 2-deoxyglucose, they were able to demonstrate that the
cells’ rhythm was passed on from one generation of cells to the next.
They then transplantated the cells from the immortal cell line into
rats that had received suprachiasmatic lesions and restored the circadian
rhythm in the rats. Thus, the rhythmic behavior of the suprachiasmatic
neurons is immortal in that it is passed from one hamster generation to
the next or from one cultured suprachiasmatic neuron to the next and
from hamster to culture and visa versa.
What Ticks?
Considerable research is being directed toward determining what genes control the
ticking of the circadian clock. Because a single suprachiasmatic neuron displays a circadian
rhythm, the timing device must be in the neuron itself, possibly entailing an
450 ! CHAPTER 12
Figure 12-7
Circadian Rhythms Restored by
Neural Transplantation Adapted from
“Transplantation: A New Tool in the Analysis
of the Mammalian Hypothalamic Circadian
Pacemaker,” by M. R. Ralph and M. N. Lehman,
1991, Trends in Neurosciences, 14, p. 363.
0
0
0
24
24
24
(A) Normal
(B) Suprachiasmatic lesion
(C) Suprachiasmatic transplant
Time (hours)
Time (hours)
Time (hours)
Normal free-running rhythm in
constant darkness
Absence of circadian rhythm
in a light–dark environment
Normal free-running rhythm in
constant darkness restored by
transplant
CH12.qxd 2/7/05 2:58 PM Page 450
ls
le
ll
increase and decrease of one or more proteins made by the cell. Just as the back-andforth
swing of a pendulum makes a grandfather clock tick, the increase and decrease
in the amount of the protein makes the cell tick once each day.
According to this notion, a protein is made until it crests at a certain level, at
which point it inhibits its own production; when its level falls to a critical point, production
again rises. In turn, the electrical activity of the cell is linked to protein oscillation,
allowing the cell to control other cells during a part of the oscillation. (The
actual way that the oscillation is produced is a little more complex than this description
suggests.)
Findings from studies on mutant and knockout mice suggest that at least a half
dozen genes and the proteins produced by them form two interlocking loops to produce
the circadian rhythm of suprachiasmatic cells in mammals (Coogan & Piggins,
2004). Although the mechanism is not fully worked out, the excitation of suprachiasmatic
cells through the retinohypothalamic pathway can presumably degrade one of
the proteins to entrain the sequence of biochemical steps in the interlocking loops.
Pacemaking Circadian Rhythms
The suprachiasmatic nucleus is of itself not responsible for directly producing rhythmic
behavior. For example, after the SCN has been damaged, the behavioral activities
of drinking and eating and of sleeping and wakefulness still occur. They no longer
occur at appropriate times, however.
One proposal for how the SCN controls behavioral rhythms is illustrated in Figure
12-8. In this model, light entrains the SCN, and the pacemaker in turn drives a
number of “slave” oscillators. Each slave oscillator is responsible for the rhythmic occurrence
of one behavior. In other words, drinking and eating, body temperature, and
sleeping and waking are each produced by a separate slave oscillator.
The signal, or “chime,” that synchronizes slave oscillators may be both hormonal
and a neurotransmitter signal from axons of suprachiasmatic neurons. Evidence that a
hormone takes part comes from findings that transplanted suprachiasmatic neurons
still send the signal, although they do not make axonal connections with distant neurons.
Because motor-activity cycles can be restored by transplants in rodents, activity
may be driven hormonally. As “Synchronizing Biorhythms at the Molecular Level” explains,
a protein made and secreted by suprachiasmatic cells may be one chime. Other
rhythms may be driven by neural connections to slave oscillators.
Shortly,we will consider some of the events of sleeping and waking behavior and the
neural mechanisms that control them. Understanding circadian rhythms is important
WHY DO WE SLEEP AND DREAM? ! 451
1 2 3
Days
Light
Eye
photoreceptors
Entrainment
pathways
Pacemaker
Output
pathways
Slave oscillators Circadian functions
Motor activity
Eating
Body temperature
Light information from
eye photoreceptors…
…entrains the suprachiasmatic
nucleus pacemaker.
The pacemaker has a
rhythm that drives
“slave” oscillators, …
…which in turn control
functions that exhibit
circadian activity.
Retina
Figure 12-8
Organization of the Circadian Timing
System
CH12.qxd 2/7/05 2:58 PM Page 451
ls
le
ll
for understanding sleeping and waking. If the circadian pacemaker function of the SCN
is disrupted, sleep is disrupted.Consequently, some sleep disorders may be due not to the
mechanisms that control sleep but to a malfunction of the pacemaker.
Pacemaking Circannual Rhythms
The suprachiasmatic nucleus not only controls daily rhythms, it can also control circannual
rhythms. Russel Reiter (1980) illustrated this form of pacemaking in hamsters.
Hamsters are summertime, or long-day, breeders. As the days lengthen in springtime,
the gonads of male hamsters grow and release hormones that stimulate the males’ sexual
behavior. As the days shorten in the winter, the gonads shrink, the amount of the
452 ! CHAPTER 12
Synchronizing Biorhythms at the Molecular Level
Focus on New Research
To be effective, a biological clock needs an input signal that
tracks light changes during cycles of day and night, the oscillator
for timekeeping, and an output signal that can drive
slave oscillators. As you know, the input signal is carried by
the retinohypothalamic pathway, which projects from special
retinal receptors through ganglion neurons. These neurons
have excitatory glutaminergic synapses onto the cells of
the suprachiasmatic nucleus.
In the rodent, this pathway stimulates SCN neurons to
be active during the light part of the cycle. Findings from molecular-
genetic studies into the clockwork show that a number
of genes and their protein products take part in timing
the SCN oscillation. The genes Period and Cryptochrome encode
the oscillator.
Period and Cryptochrome are switched on by the proteins
Clock and Bmal and switched off by the protein complex
of Per and Cry so that gene turn-on follows gene turn-off
in an inexorable daily loop. Mutations either in Period or
Cryptochrome can lead to inherited sleep disorders. In addition,
investigations into the sleeping habits of people suggest
that modifications in genes that produce other proteins, such
as Clock, can influence whether an individual will be “early
to bed and early to rise” or “late to bed and late to rise”
(Archer et al., 2003).
How does the SCN signal the slave oscillators in the rest
of the brain to synchronize body temperature, metabolic activity,
growth-hormone release, sleep activity, and so forth,
with the light–dark cycle? A puz zling anatomical feature of
SCN neurons is that they do not all form major pathways
from the SCN to other nuclei in the brain over which the
“chime” that signals circadian activity can be sent. In addition,
if transplanted SCN neurons are grafted in place of a
damaged SCN, the new SCN chime is sufficient to signal
other brain regions even though the grafted neurons do not
form axons that project to the SCN’s usual targets.
These findings suggest that one chime from the SCN is
chemical. Michelle Cheng and her colleagues (2003) described
six lines of evidence suggesting that a protein called
prokineticin 2 (PK2) may be a SCN chime in mice:
1. PK2 levels are approximately 50 times as high in SCN
neurons during the light (inactive) part of the cycle as in
the dark (active) part of the cycle.
2. The production of PK2 is controlled by Clock genes.
3. In mice with deficient Clock genes, the expression of
PK2 in SCN neurons was extremely low.
4. After mice maintained in constant darkness were exposed
to light, the level of PK2 increased rapidly.
5. Receptors for PK2 are found in a number of brain regions
that that are thought to serve as slave oscillators.
6. The administration of PK2 by injection into the brain by
a cannula during the dark period of the circadian cycle
resulted in a suppression of the expected increase of locomotor
activity displayed by the mice.
CH12.qxd 2/7/05 2:58 PM Page 452
ls
le
ll
hormones produced by the gonads decreases, and the males stop being interested in
sexual behavior.
During the dark phase of the day–night cycle, the pineal gland secretes the hormone
melatonin. This hormone influences daily and seasonal biorhythms. Figure 12-9
shows that, when a hamster’s melatonin level is low, gonads enlarge and, when it is
high, gonads shrink. The control that the pineal gland exerts over the gonads is in turn
controlled by the suprachiasmatic nucleus. Through a rather indirect pathway, the SCN
drives the pineal gland as a slave oscillator.
During the daylight period of the circadian cycle, the SCN inhibits melatonin secretion
by the pineal gland. Thus, as the days become shorter, the period of inhibition
becomes shorter and thus the period in which melatonin is released becomes longer.
When the period of daylight is shorter than 12 hours, melatonin release becomes sufficiently
long to inhibit the hamster’s gonads so that they shrink.
Melatonin also influences the testes of animals that are short-day breeders, such as
sheep and deer, which mate in the fall and early winter. Its effect on reproductive behavior
in these species is the reverse of that in the hamster: their reproductive activities
begin as melatonin release increases.
The origins of many biorhythms are not yet understood. In his book titled Biological
Clocks in Medicine and Psychiatry, Curt Richter (1965) summarized a lifetime of
recording various normal and abnormal rhythms. Richter recorded rhythmic activity
in many bodily functions—including body temperature, hormone levels, eating, and
drinking.
Richter hypothesized that many physical and behavioral disorders might be caused
by “shocks,” either physical or environmental, that upset the timing of biological clocks.
For example, the record of psychotic attacks suffered by English writer Mary Lamb, illustrated
in Figure 12-10, is one of many rhythmic records that Richter thought represented
the action of an abnormally functioning biological clock.
WHY DO WE SLEEP AND DREAM? ! 453
Light Dark Light Dark Gonadal size
Spring
Fall
Gonadal size
Less
testosterone More
testosterone
Melatonin
level
Winter Summer
Melatonin
level
As daylight lengthens in summer,
melatonin production decreases, allowing
gonads to grow and to increase hormone
production to stimulate sexual behavior.
As daylight shortens in winter,
increases in melatonin levels
cause gonads to shrink,
decreasing hormone
production and sexual interest.
Age
29 31 33 35 37 39 41 43 45 47 49 51 53 55 57 59 61 63 65 67 69 71 73 75 77 79 81 83
Figure 12-10
Dysfunctional Clock? Attacks of
mental illness displayed by the English
writer Mary Lamb through her adult
life appear to have had a cyclical
component. Such observations would be
difficult to obtain today, because the
drugs used to treat psychiatric disorders
can mask abnormal biorhythms. Adapted
from Biological Clocks in Medicine and Psychiatry
(p. 92), by C. P. Richter, 1965, Springfield, IL:
Charles C Thomas.
Figure 12-9
A Hamster’s Circannual Pacemaker
Adapted from “The Pineal and Its Hormones
in the Control of Reproduction in Mammals,”
by R. J. Reiter, 1980, Endocrinology Review, 1,
p. 120.
Melatonin. Hormone secreted by the
pineal gland during the dark phase of the
day–night cycle; influences daily and
seasonal biorhythms.
CH12.qxd 2/7/05 2:58 PM Page 453
ls
le
ll
454 ! CHAPTER 12
SLEEP STAGES AND DREAMING
Most of us are awake during the day and asleep at night. Both behavioral states
are more complex than our daily experiences suggest.Waking behavior encompasses
some periods in which we are relatively still, other periods in which we are still but
mentally active, and still other periods in which we are physically active. Our sleeping
behavior is similarly variable in that it consists of periods of resting, napping,
long bouts of sleep, and various sleep-related events including snoring, dreaming,
thrashing about, and even sleepwalking. In this section, we describe some of the behavioral
events of waking and sleeping and some of the neural processes that underlie
them.
Measuring How Long We Sleep
A crude measure of sleeping and waking behavior is the self-report; that is, people
record in a diary when they wake and when they retire to sleep. These diaries show considerable
variation in sleep–waking behavior. People sleep more when they are young
than when they are old.Most people sleep about 7 to 8 hours per night, but some people
sleep much more or less than that, even as little as 1 hour each day.
Some people nap for a brief period in the daytime, and others never nap. Benjamin
Franklin is credited with the aphorism, “Early to bed and early to rise makes a man
healthy, wealthy, and wise,” but measures of sleep behavior indicate that the correlation
that Franklin made does not actually exist. Apparently, variations in sleeping times are
quite normal.
Measuring Sleep in the Laboratory
In contrast with self-reports, laboratory sleep studies allow researchers to record
physiological changes associated with sleep. The electrical activity in the brain and
body is measured with a polygraph, as described in Chapter 4. Figure 12-11 illustrates
a typical polygraph setup in a sleep laboratory and some commonly used
measures.
Electrodes are pasted onto a number of standard locations on the skull’s surface
for an electroencephalogram (EEG), a record of brain-wave activity; onto muscles of
In Review .
A number of nuclei in the brain serve as diurnal biological clocks, including the suprachiasmatic
nucleus of the hypothalamus and the pineal gland of the thalamus. Cues from
the environment called Zeitgebers entrain the suprachiasmatic nucleus so that it can
accurately control daily rhythms. Damage to the suprachiasmatic nucleus disrupts the
rhythm of daily behaviors. We know that the pacemaking produced by the suprachiasmatic
nucleus is a product of its cells because, if removed and cultured in a dish,
the cells continue their rhythmic behavior and even pass on their rhythms to offspring
cells cultured in a dish. If such immortal cells are transplanted back into a brain from
which the suprachiasmatic nucleus has been removed, they restore the animal’s rhythmic
behavior.
CH12.qxd 2/7/05 2:58 PM Page 454
ls
le
ll
the neck for an electromyogram (EMG), a record of muscle activity;
and above the eyes for an electrooculogram (EOG), a record of eye movements.A thermometer
also may be used to measure body temperature. Together, these measures
provide a comprehensive physiological measure of sleeping–waking states.
The EEG recording traces distinct patterns of brain-wave activity and is the primary
measure of sleep states. The neocortex generates rhythmic patterns from states
categorized as awake, drowsy, sleeping, and dreaming.
Waking State When a person is awake, the EEG pattern consists of small-amplitude
(height) waves with a fast frequency. This pattern, the beta rhythm (b), is also called
fast activity, activated EEG, or waking EEG. The waves of the beta rhythm have a frequency
(repetition period) ranging from 15 to 30 Hz (times per second).
Drowsy State When a person becomes drowsy, the fastwave
activity of the neocortex slows down and deepens
concurrently. The amplitude of the waves increases, and
the frequency of the waves becomes slower. When subjects
relax and close their eyes, they may produce the
alpha rhythm, large, extremely regular brain waves that
have a frequency ranging from 7 to 11 Hz (review Figure
4-25).
In humans, you may recall, alpha rhythms are generated
in the region of the visual cortex at the back of the
brain, and they abruptly stop if a relaxed person is disturbed
or opens his or her eyes. Recall, too, that not everyone
displays alpha rhythms, and some people display them
much better than others.
WHY DO WE SLEEP AND DREAM? ! 455
Figure 12-11
Setup in a Sleep Laboratory Electronic equipment records
readouts from the electrodes attached to the sleeping subject.
(A) Electroencephalogram made from a point on the skull relative to a
neutral point on the ear. (B) Electromyogram made between two muscles,
such as those on the chin and throat. (C) Electrooculogram made between
the eye and a neutral point on the ear.
Beta rhythm. Fast brain-wave activity
pattern associated with a waking EEG.
Excited
Relaxed, eyes closed
1
Time (s)
2 3
Deep sleep
(C) Electrooculogram (EOG)
(B) Electromyogram (EMG)
(A) Electroencephalogram (EEG)
Hank Morgan/Rainbow
CH12.qxd 2/7/05 2:58 PM Page 455
ls
le
ll
Sleeping State As subjects enter deep sleep, they produce yet slower,wider EEG waves
known as delta rhythms (d), also known as slow-wave activity or resting activity.Delta
rhythm waves have a frequency of 1 to 3 Hz. The slowing of brain-wave activity is also
associated with the loss of consciousness that characterizes sleep.
Dreaming State Sleep consists of periods during which a sleeper is relatively still and
periods when the mouth, fingers, and toes twitch. These opposite aspects of sleep are
readily observable in household pets and bed partners. In 1955, Eugene Aserinsky and
Nathaniel Kleitman (Lamberg, 2003), working at the University of Chicago, observed
that the twitching periods are also associated with rapid eye movements (REMs).
Rapid eye movements coincide with distinct brain-wave patterns recorded on the
EEG that suggest that the subject is awake, with the eyes flickering back and forth behind
the sleeper’s closed eyelids (see Dement, 1972). By accumulating and analyzing
REMs recorded on EEGs, the Chicago investigators were the first to identify REM sleep,
the fast-wave pattern displayed by the neocortical EEG record. The EEG record suggested
that dreaming subjects were awake, even though Aserinsky and Kleitman confirmed
that the subjects really were asleep.
A Typical Night’s Sleep
Because fast-wave activity is associated with REM sleep, the phase of sleep associated
with delta rhythms on the EEG recording is called NREM (for non-REM) sleep and
sometimes slow-wave sleep. With this distinction between sleep phases in mind, we
now turn to comparing the EEG patterns associated with the course of a typical night’s
sleep.
Figure 12-12A displays the EEG patterns associated with waking, sleeping, and
dreaming. Non-REM sleep is divided into four stages on the basis of EEG records. Notice
that the main change characterizing these stages is that brain waves become larger
and slower in a progression from stage 1 sleep through stage 4 sleep.
The designation of these stages assumes that the sleeper moves from relatively shallow
sleep in stage 1 to deeper sleep in stage 4. Self-reports of subjects who are awakened
from sleep at different times suggest that stage 4 is the deepest sleep because subjects act
groggy when disturbed in these periods. A remarkable feature of sleep is that the EEGs
456 ! CHAPTER 12
Delta rhythm. Slow brain-wave activity
pattern associated with deep sleep.
REM sleep. Fast brain-wave pattern
displayed by the neocortical EEG record
during sleep.
Non-REM sleep. Slow-wave sleep
associated with delta rhythms.
Slow-wave sleep. NREM sleep.
(B) Sleep
Depth of sleep
REM
REM
REM
REM
REM
Time
(hr)
7
3
2
1
6
5
4
NREM
dominant
REM
dominant
Stage 4
Stage 2
Stage 1
REM
(A) EEG
Awake
Stage 3
Figure 12-12
Sleep Recording and Revelations
(A) Electroencephalograph patterns
associated with waking, with the four
NREM sleep stages, and with REM sleep.
(B) In a typical night’s sleep, a person
undergoes a number of sleep-state
changes, roughly in 90-min periods.
Non-REM sleep dominates the early
sleep periods, and REM sleep dominates
later sleep. The duration of each stage
of sleep is indicated by the thickness of
each bar, which is color-coded to the
corresponding stage in part A, with the
dark purple REM sleep bars labeled. The
depth of each stage is graphed as the
relative length of the bar. Adapted from
“Sleep and Dreaming,” by D. D. Kelley, in E. R.
Kandel, J. H. Schwartz, and T. M. Jessell (Eds.),
Principles of Neuroscience, 1991, New York:
Elsevier, p. 794.
Visit the section on the EEG in the
Research Methods module of your CD to
learn more about the EEG and the stages
of sleep and waking.
CH12.qxd 2/7/05 2:58 PM Page 456
ls
le
ll
of waking, of stage 1 sleep, and of REM sleep are similar. In all three conditions, the
brain waves have an active, beta-rhythm pattern. Other unique physiological events
occur in REM. The oculograph, for example, indicates that the subject’s eyes are moving,
revealing that it is recording REM sleep.
On the basis of a record of brain activity from one subject during a typical night’s
sleep, Figure 12-12B graphs when these different sleep stages actually occur and how
long they last in the course of a night’s sleep. Notice that the depth of sleep changes
several times. The subject cycles through the four stages of NREM sleep and then enters
REM sleep. This NREM–REM sequence lasts approximately 90 min and is typically
repeated about four times in the course of the sleep period.
You can see by the labels indicating REM sleep in Figure 12-12B that the durations
of the different sleep stages roughly divide the sleep period into two parts, the first
dominated by NREM sleep and the second dominated by REM sleep. Body temperature
is lowest (about 1.5 degrees below a normal temperature of 37.7°C) during the
first part of the sleep period and rises during the second part.When subjects are awakened
in REM sleep, they appear more alert and attentive than they are when awakened
at other times, and they report that they have been vividly dreaming.
Findings from studies of sleep in the laboratory confirm that the sleep of individual
people is highly variable; they also confirm that REM sleep takes up a substantial
proportion of sleep time. Adults who typically sleep about 8 hours spend
about two of those hours in REM sleep. A person’s
REM durations may also vary at different
times of life. Periods of REM sleep increase during
growth spurts, in conjunction with physical
exertion, and, for women, during pregnancy.
The time spent in REM sleep also changes
dramatically over the life span. As is illustrated in
Figure 12-13, most people sleep less as they grow
older. Furthermore, in the first 2 years of life, REM
sleep makes up nearly half of sleep time, but it declines
proportionately until, in middle age, it constitutes
little more than 10 percent of sleep time.
Non-REM Sleep
Although many people may think that sleep is an inactive period, a remarkable range of
activities take place during sleep (see “Restless Legs Syndrome”). During NREM sleep,
body temperature declines, heart rate decreases, blood flow decreases, we perspire and
lose body weight owing to water loss, and our levels of growth hormone increase.
It was once thought that we do not dream during NREM sleep, but findings from
recent studies show that, when subjects are aroused from NREM sleep, they do report
dreams. These dreams lack the vividness of dreams reported by subjects aroused from
REM sleep, however. Non-REM sleep is also the time during which we toss and turn
in bed, pull on the covers, and engage in other movements. If we talk in our sleep, we
will do so during NREM sleep. If we make flailing movements of the limbs, such as
banging with an arm or kicking with a foot, we will usually do so in NREM sleep.
Some people even get up and walk while asleep, and this “sleepwalking” takes place
in NREM sleep. Children may experience brief, very frightening dreams called night
terrors, which also occur in NREM sleep. All these conditions are inconsistent with a
period that is often described as quiet and inactive.
WHY DO WE SLEEP AND DREAM? ! 457
To learn more about night terrors, visit
the Web site at www.worthpublishers.
com/kolb/chapter12
Figure 12-13
Sleeping and Waking
over the Life Span The
amount of time that
humans spend sleeping
decreases with age. The
amount of REM sleep is
especially high in the first
few years of life. Adapted
from “Ontogenetic Development
of the Human Sleep-Dream
Cycle,” by H. P. Roffward,
J. Muzio, and W. C. Dement
(1966), Science, 152.
1 10 30 60 90
Age (years)
0
8
10
12
16
24
Hours
Waking
REM
sleep
NREM
sleep
CH12.qxd 2/7/05 2:58 PM Page 457
ls
le
ll
REM Sleep and Dreaming
REM sleep is no less exciting and remarkable than NREM sleep. During REM sleep,
our eyes move and our toes, fingers, and mouths twitch, and males have penile erections.
Still, we are paralyzed, as indicated by atonia (no tone), the absence of muscle
tone due to the inhibition of motor neurons. Atonia is recorded on an EMG by the absence
of muscle activity.
458 ! CHAPTER 12
Restless Legs Syndrome
Focus on Disorders
I’ve always been a fairly untalented sleeper. Even as
a child, it would take me some time to fall asleep,
and I would often roll around searching for a comfortable
position before going under. But my real
difficulties with sleeping did not manifest themselves
until early adulthood. By that time, my father
had been diagnosed with Restless Legs Syndrome
(RLS) and I was suffering the same symptoms.
Initially, my symptoms consisted of a mild tingling
in my legs. It caused me to be fidgety and made
it hard to fall asleep. Eventually, I went through a
number of days without much sleep and reached a
point where I simply could not function. I went to a
doctor who prescribed a small course of sleeping
medication (a benzodiazepine). I was able to get
good sleep and my sleep cycle seemed to get back on
track. Over the next decade I had periodic bouts of
tingling in my legs which caused me to be fidgety and
interfered with sleep. As time passed, the bouts occurred
with increasing frequency and the symptoms
became more noticeable and uncomfortable. I would
simply suffer through these bouts, sleeping poorly
and paying the consequences, or I would seek medical
help. Being a student, I did not have a regular
doctor. Unfortunately, most physicians I met did not
know about RLS, and thought I was “drug seeking” or
merely stressed out. I received a variety of patronizing
responses and found these experiences insulting
and demeaning. It would have been easy to give up
and try to deal with it on my own, but because of my
father’s diagnosis, I knew the true source of the problem
and was determined to get help.
When I took my current position, I started seeing
a doctor on a regular basis. By this time, my
sleep was being seriously affected by RLS. The sensations
in my legs were something like a combination
of an ache in my muscles (much like one gets
after exercising) and an electrical, tingling sensation.
They would be briefly relieved with movement,
such as stretching, rubbing, contracting my
muscles, or changing position, but would return
within seconds. In fact, my wife says my cycle is
about 13 to 15 seconds between movements. I do
this either when awake or during sleep. Trying not
to move greatly increases the discomfort—much
like trying to not scratch a very bad itch. The symptoms
get worse in the evening and at night. Most
nights, I have trouble falling asleep. Other nights, I
Bill Aron/PhotoEdit
Atonia. No tone; condition of complete
muscle inactivity produced by the
inhibition of motor neurons.
CH12.qxd 2/7/05 2:58 PM Page 458
ls
le
ll
You can get an idea of what REM sleep is like by observing a cat or dog.At the onset
of REM sleep, the animal usually subsides into a sprawled posture as the paralysis of its
muscles sets in. Figure 12-14 illustrates the sleep postures of a horse. Horses can sleep
while standing up by locking their knee joints, and they can sleep while lying down with
their heads held slightly up.At these times, they are in NREM sleep; when they are completely
sprawled out, they are in REM sleep.
WHY DO WE SLEEP AND DREAM? ! 459
wake up after an hour or so and then have trouble
going back under.
Now my doctor takes me seriously. We exchanged
research articles and thoroughly discussed
treatment options. As part of this process, my wife
met with my physician to relate her experiences. I
was stunned to learn how severely my RLS was interfering
with her sleep. I think she was being compassionate
and not complaining so I wouldn’t feel
any more upset than I already was about my sleeping
difficulties. My doctor started me on a regular
course of sleeping medication (again, a benzodiazepine)
and encouraged me to stay on it. This was
a life-changing event. For the first time in my adult
life, I was getting good sleep on a regular basis. My
wife and I also got separate beds.
On the down side, RLS is a chronic and progressive
condition. Over the years I’ve had to slowly
increase dosages, switch to new medicines, and am
now on two medications. I have always communicated
openly and honestly with my physician, and
we have worked together to monitor issues such as
tolerance and medication dosages. I have gone for
consultation with a neurologist experienced with
RLS. I still get good sleep, but I have had to make
many significant adjustments.
I am very up front about the fact that I have
RLS. In fact, whenever I teach the topic of sleep and
sleep disorders in my brain and behavior classes, I
always make some time to talk about my experiences
with RLS. Occasionally, students approach
me with their own difficulties, and I try to provide
them with information and resources.
—Stuart Hall, Ph.D., University of Montana
Restless legs syndrome (RLS) is a sleep disorder in which a
person experiences unpleasant sensations in the legs described
as creeping, crawling, tingling, pulling, or pain. The
sensations are usually in the calf area but may be felt anywhere
from the thigh to the ankle. One or both legs may be
affected; for some people, the sensations are also felt in the
arms.
People with RLS describe an irresistible urge to move
the legs when the sensations occur. Many people with RLS
have a related sleep disorder called periodic limb movement
in sleep (PLMS). It is characterized by involuntary
jerking or bending leg movements in sleep that typically
occur every 10 to 60 s. Some people experience hundreds
of such movements per night, which can wake them, disturb
their sleep, and annoy bed partners. People with these
disorders get less sleep at night and may feel sleepy during
the day.
These symptoms affect both sexes, and symptoms
can begin at any time but are more severe among older
people. Young people who experience symptoms are sometimes
thought to have “growing pains” or may be considered
hyperactive because they cannot easily sit still in
school.
There is no laboratory test for these disorders, and a
doctor cannot detect anything abnormal in a physical examination.
The disorder may also be of central nervous system
origin because the syndrome has been reported in a
patient who had no legs (Estivill et al., 2004). In mild cases,
massage, exercise, stretching, and hot baths may be helpful.
For more severe cases, patients can restrict their intake
of caffeine, take benzodiazepines to help them get to sleep,
and take L-dopa, a drug that is also used to treat Parkinson’s
disease. These treatments reduce symptoms, but at present
there is no cure for the condition.
CH12.qxd 2/7/05 2:58 PM Page 459
ls
le
ll
During REM sleep,mammals’ limbs twitch visibly, and, if you look carefully at the
face of a dog or cat, you will also see the skin of the snout twitch and the eyes move behind
the eyelids. It might seem strange that an animal that is paralyzed can make small
twitching movements, but the neural pathways that mediate these twitches are presumably
spared the paralysis. One explanation for the twitching movements of the
eyes, face, and distal parts of the limbs is that such movements may help to maintain
blood flow in those parts of the body.
An additional change resulting from atonia during REM sleep is that mechanisms
that regulate body temperature stop working and body temperature moves toward
room temperature. The sleeper may wake up from REM sleep feeling cold or hot, depending
on the temperature of the room.
The most remarkable aspect of REM sleep—dreaming—was discovered by William
Dement and Nathaniel Kleitman in 1957 (Dement, 1972). When subjects were
awakened from REM sleep, they reported that they had been having vivid dreams. In
contrast, subjects aroused from NREM sleep were much less likely to report that they
had been dreaming, and the dreams that they did report were much less vivid. The
technique of electrical recording from a sleeping subject in a sleep laboratory made it
possible to subject dreams to experimental analysis, and such studies provided some
objective answers to a number of interesting questions concerning dreaming.
The first question that studies of dreaming answered was, How often do people
dream? Reports by people on their dreaming behavior had previously suggested that
dreaming was quite variable, with some people reporting that they dreamed frequently
and others reporting that they never dreamed.Waking subjects up in periods of REM
showed that everyone dreams, that they dream a number of times each night, and that
dreams last longer as a sleep session progresses. Those who claimed not to dream were
presumably forgetting their dreams. Perhaps people forget their dreams because they
do not wake up in the course of a dream or immediately afterward, thus allowing subsequent
NREM sleep activity to erase the memory of the dream.
Another interesting question that objective measures answered was, How long do
dreams last? There had been suggestions that dreams last but an instant. By waking
people up at different intervals after the onset of a REM period and matching the reported
dream content to the previous duration of REM sleep, researchers were able to
show that dreams appear to take place in real time. That is, an action that a person performed
in a dream lasts about as long as it would take to perform while awake.
What We Dream About
The study of dreaming in sleep laboratories also allows researchers to study some of
the questions that have always intrigued people.Why do we dream? What do we dream
about? What do dreams mean?
460 ! CHAPTER 12
Figure 12-14
Nap Time Horses usually seek an
open, sunny area in which to take brief
periods of sleep. Ian’s horse, Lady Jones,
illustrates three sleep postures. At left,
she displays NREM sleep, standing with
legs locked and head down, and, at
center, she displays NREM sleep lying
down with head up. At right, she is in
REM sleep, in which all postural and
muscle tone is lost.
Courtesty of Ian Wishaw
Courtesty of Ian Wishaw
Courtesty of Ian Wishaw
CH12.qxd 2/7/05 2:58 PM Page 460
ls
le
ll
Past explanations of dreaming have ranged from messages from the gods to indigestion.
The first modern treatment of dreams was described by the founder of psychoanalysis,
Sigmund Freud, in The Interpretation of Dreams, published in 1900. Freud
reviewed the early literature on dreams, described a methodology for studying them,
and provided a theory to explain their meaning.We briefly consider Freud’s theory because
it remains popular in psychoanalysis and in the arts.
Freud suggested that the function of dreams was the symbolic fulfillment of unconscious
wishes. His theory of personality was that people have both a conscious and
an unconscious. Freud proposed that the unconscious contains unacknowledged desires
and wishes, which are sexual. He further proposed that dreams have two levels of
meaning. The manifest content of a dream consists of a series of often bizarre images
and actions that are only loosely connected. The latent content of the dream contains
its true meaning, which, when interpreted by a psychoanalyst, provides a coherent account
of the dreamer’s unconscious wishes.
Freud provided a method for interpreting manifest symbols and reconstructing
the latent content of dreams. For example, he pointed out that a dream usually begins
with an incident from the previous day, incorporates childhood experiences, and includes
ongoing unfulfilled wishes.He also identified a number of types of dreams, such
as those that deal with childhood events, anxiety, and wish fulfillment. The content of
the dream was important to Freud and other psychoanalysts in clinical practice because,
when interpreted, dreams served as a source of insight into a patient’s problems.
Other psychoanalysts, unhappy with Freud’s emphasis on sexual desire, developed
their own methods of interpretation. Psychoanalyst Carl Jung, a contemporary of Freud,
proposed that the symbolism of dreams signifies distant human memories encoded in
the brain but long since lost to conscious awareness. Jung proposed that dreams allow a
dreamer to relive the history of the human race, our “collective unconscious.” As more
theories of dream interpretation developed, their central weakness became apparent: it
was difficult, if not impossible, to know which interpretation was correct.
The dream research of Freud and his contemporaries was impeded by their reliance
on a subject’s memory of a dream and by the fact that many of their subjects were patients.
This situation unquestionably resulted in the selection of the unusual by both the
patient and the analyst. Now that researchers study dreams more objectively by waking
subjects and questioning them, one might think that the meaning of dreams might be
better understood. Certainly, knowledge of the content of dreams has improved.
Research suggests that most dreams are related to events that happened quite recently
and concern ongoing problems. Colors of objects, symbols, and emotional content
most often relate to events taking place in a person’s recent waking period. Calvin
Hall and his colleagues (1982) documented more than 10,000 dreams of normal people
and found that more than 64 percent are associated with sadness, anxiety, or anger.
Only about 18 percent are happy. Hostile acts against the dreamer outnumber friendly
acts by more than two to one. Surprisingly, in regard to Freud’s theory, only about 1
percent of dreams include sexual feelings or acts.
Contemporary dream hypotheses fall into two groups: those stating that dreams
have no meaning and those stating that dreams are meaningful. The views of a major
proponents of each position are described next.
J. Allan Hobson (2004), a scientist who has devoted his life to the scientific study of
sleep and dreaming, proposed in his activation–synthesis hypothesis that, during a
dream, the cortex is bombarded by signals from the brainstem, and these signals produce
the pattern of waking (or activated) EEG. The cortex, in response to this excitation, generates
images, actions, and emotion from personal memory stores. In the absence of external
verification, these dream events are fragmented and bizarre and reveal nothing
more than that the cortex has been activated. Furthermore, Hobson proposed, on the
WHY DO WE SLEEP AND DREAM? ! 461
Click on the Web site to investigate
more about research on dreams at
www.worthpublishers.com/kolb/
chapter12
CH12.qxd 2/7/05 2:58 PM Page 461
ls
le
ll
462 ! CHAPTER 12
basis of PET-imaging results, that part of the frontal cortex is less active in dreaming than
in waking. Because the frontal cortex controls working memory,memory for events that
have just happened, and attention, the dreamer has difficulty both in remembering and
linking dream events as they take place and in deciding what dream events should follow
other events.
According to Hobson’s hypothesis, dreams are nevertheless personal in that memories
and experiences are activated, but they have no meaning. So, for example, the
following dream, with its bizarre, delusional, and fragmented elements, would be representative
of images that are synthesized to accompany brain activation. According to
Hobson’s hypothesis, any meaning that the dream might seem to have is created by the
dreamer after the fact, as was perhaps done by the middle-aged dreamer who recounted
this dream:
I found myself walking in a jungle. Everything was green and fresh and I felt
refreshed and content. After some time I encountered a girl whom I did not
know. The most remarkable thing about her was her eyes, which had an almost
gold color. I was really struck by her eyes not only because of their
unique color but also because of their expression. I tried to make out other details
of her face and body but her eyes were so dominating that was all I could
see. Eventually, however, I noticed that she was dressed in a white robe and was
standing very still with her hands at her side. I then noticed that she was in a
compound with wire around it. I became concerned that she was a prisoner.
Soon, I noticed other people dressed in white robes and they were also standing
still or walking slowly without swinging their arms. It was really apparent
that they were all prisoners. At this time I was standing by the fence that enclosed
them, and I was starting to feel more concerned. Suddenly it dawned
on me that I was in the compound and when I looked down at myself I found
that I was dressed in a white robe as well. I remember that I suddenly became
quite frightened and woke up when I realized that I was exactly like everyone
else. The reason that I remembered this dream is the very striking way in
which my emotions seemed to be going from contentment, to concern, to fear
as the dream progressed. I think that this dream reflected my desire in the
1970s to maintain my individuality. (Recounted by A.W.)
Anttio Revonsuo (2000) of Finland agrees with Hobson about the content of
dreams but uses content analysis to argue that dreams are biologically important in that
they lead to enhanced performance in dealing with threatening life events. In his evolutionary
hypothesis of dreams, Revonsuo argues that this enhanced performance would
have been especially important for early humans,whose environment included frequent
dangerous events that constituted extreme threats to their reproductive success. He
notes that dreams are highly organized and are significantly biased toward threatening
images (as, e.g., in the preceding dream). People seldom dream about reading, writing,
and calculating, even though these behaviors may occupy much of their day.
The threatening events of dreams are the same ones that are threatening in real life
(Figure 12-15). For example, animals and strange men who could be characterized as
“enemies” figure prominently in dreams. Revonsuo cites overwhelming evidence that
dream content incorporates the current emotional problems of the dreamer. He also
reviews evidence to suggest that depressed dreamers who dream about their focal problems
are better adjusted than those who do not.
Revonsuo also notes that recurrent dreams and nightmares generally begin in childhood,
when a person is most vulnerable, and are associated with anxiety, threats, and
pursuit. In them, the dreamer is usually watching, hiding, or running away. Revonsuo
CH12.qxd 2/7/05 2:58 PM Page 462
ls
le
ll
therefore proposes that the experience of dealing with threats in dreams is adaptive because
it can be applied to dealing with real-life threats. To illustrate, a student provided
the following account of a dream from childhood that she had dreamed subsequently a
few times:
When I was five years old, I had a dream that at the time frightened me but
that I now find somewhat amusing. It took place in the skating rink of my
small hometown. There was no ice in the rink, but instead the floor consisted
of sod. The women, my mother included, were working in the concession
booth, and the men were in the arena, dressed in their work clothes. I was
among the children of the town who were lined up in the lobby of the rink.
None of the children, including myself, knew why we were lined up. The adults
were summoning the children two at a time. I decided to take a peek through
the window, and this is what I saw. There was a large circus-ride type metal
chair that was connected to a pulley, which would raise the chair to about 20
feet into the air. The seat would be lowered and two children at a time would
be placed in it. A noose was then placed around the neck of each and the chair
was again raised. Once the chair reached its greatest height, the bottom would
drop out of the chair and the children would be hanged (I did not see this but
I thought that is what happened to them). At this point, I turned to a friend
and said, “Here,Ursula, you can go ahead of me” and I went to my mother and
told her what was going on. She smiled as if I were just being difficult and told
me that I was to get back in the line. At this point I thought, “Forget it,” and I
found a place to hide underneath the big wooden bleachers in the lobby. It was
dark and I could hear everyone out looking for me. (Recounted by N.W.)
When asked what she thought this dream meant, this student said that she really
did not know. When told that it could be an anxiety dream, something common in
children, that might represent an activity that she considered stressful, such as competing
in figure skating and failing, she said that she did not think that was it. She volunteered,
however, that her community’s skating rink was natural ice and that it was
bitterly cold whenever there was enough ice to go skating.When she had to skate, her
feet got cold and her mother almost had to lift her up and drag her out onto the ice.
Being dropped out of the chair may have been a symbolic representation of being
pushed out onto the ice.
Elements of the dream did represent what went on at the skating rink. Men did
prepare the ice and the women did run a concession booth, and she did resist being
sent out to skate. The recurrence of the dream could be due to the conflict that she felt
about having to do something that exposed her to the cold and her solution in hiding.
WHY DO WE SLEEP AND DREAM? ! 463
Figure 12-15
Dream Content The terrifying
visions that may persist even after
awakening from a frightening dream
are represented in this painting titled
The Night, by Swiss artist Ferdinand
Hodler. The Night, by Ferdinand Hodler
(1853–1918), oil on canvas, 116 ! 299 cm,
Kunstmuseum, Berne, Switzerland.
Erich Lessing/Art Resource
CH12.qxd 2/7/05 2:58 PM Page 463
ls
le
ll
464 ! CHAPTER 12
WHAT DOES SLEEP ACCOMPLISH?
The simplest question that we can ask about sleep is, Why do we sleep? Any satisfactory
explanation has a lot to account for. As we have seen, sleep is complex and progresses
through periodic stages.
Sleep’s rhythmic component lasts about 90 min in the course of which brain-wave
activity gradually slows and then speeds up again. In REM sleep, the sleeping brain has
a waking EEG, the motor system is paralyzed except for twitching movements, and
people have more-vivid dreams than those in other stages of sleep. An adequate theory
of sleep must account for all these phenomena. This section summarizes four theories
of why we sleep.
Sleep As a Passive Process
One of the earliest explanations views sleep as a passive process that takes place as a result
of a decrease in sensory stimulation. According to the theory, as evening approaches,
there are fewer stimuli to maintain alertness, and so sleep sets in. This theory
does not account for the complexity of sleep, nor is it supported by direct experimental
investigations.
It predicts that, if subjects are deprived of all stimulation, they will go to sleep.
Recall from Chapter 11, however, that findings from sensory-deprivation research fail
to bear out this prediction.And the results of sleep experiments reveal that, when subjects
are isolated in quiet bedrooms, they spend less, not more, time asleep. These results
do not support the idea that sleep sets in because there is nothing else to do.
The passive-process theory of sleep originally
did not consider biological rhythms as a contributing
factor to sleep, but what we now know about
biological rhythms provides some support for a
weak version of this idea. Shimon Amir and Jane
Stewart (1996) showed that initially neutral stimuli
can be conditioned to be Zeitgebers, which entrain
more-regular circadian rhythms and thus more-
In Review .
The average length of a night’s sleep is from 7 to 8 hours, but some people sleep much
less or much more. Sleep consists of two states, rapid eye movement, or REM, sleep and
non-REM, or NREM, sleep. Non-REM sleep is divided into four stages on the basis of the
EEG record. Brain waves in stage 1 sleep resemble waking patterns and REM sleep.
Those in stages 2 through 4 are characterized by progressive slowing of the EEG record.
People may have less-vivid dreams in non-REM sleep and will toss and turn. There are
about four REM sleep periods each night, with each period getting longer as sleep progresses.
REM sleep is also marked by muscle paralysis and dreams that are more vivid
than those of NREM sleep. Among the various interpretations of the function of dreams,
the activation–synthesis hypothesis suggests that they are simply a by-product of the
brain’s activity and so have no meaning, whereas the evolutionary hypothesis suggests
that dreams help people to work out solutions to threatening problems and events.
Shimon Amir Jane Stewart
Basic rest–activity cycle (BRAC).
Recurring cycle of temporal packets,
about 90-min periods in humans, during
which an animal’s level of arousal waxes
and wanes.
CH12.qxd 2/7/05 2:58 PM Page 464
ls
le
ll
regular periods of sleep. In other words, the activities in which we engage before we
sleep and after we wake up will become Zeitgebers by being associated with light–
dark changes. Therefore, exposure to darkness and quiet in the evening and to light
and other kinds of stimulation in the morning is one way of synchronizing biological
rhythms.
Sleep As a Biological Adaptation
Another explanation holds that sleep is a biologically adaptive behavior influenced by
the many ways in which a species adapts to its environment:
Sleep is designed as an energy-conserving strategy to cope with times when food is
scarce, as you learned in this chapter’s first Focus. Each animal species gathers food at
optimal times and conserves energy the rest of the time. If the nutrient value of the
food that a species eats is high, the species can spend less time foraging and more time
sleeping.
An animal’s behavior is influenced by whether the species is predator or prey. The
predator can sleep at its ease; the prey’s sleep time is reduced because it must remain
alert and ready to fight or flee at unpredictable times (Figure 12-16).
An animal that is strictly nocturnal or diurnal will likely sleep when it cannot travel
easily. Dement has stated this idea as follows: “We sleep to keep from bumping into
things in the dark.”
The sleep patterns of most animal species are consistent with the adaptive explanation.
Figure 12-17 charts the average sleep time of a number of common mammals.
Herbivores, including donkeys, horses, and cows, spend a long time collecting enough
food to sustain themselves, which reduces their sleep time. Because they are also prey,
their sleep time is further reduced as they watch for predators. Carnivores, including
domestic cats and dogs, eat nutrient-rich foods and usually consume
most of a day’s or even a week’s food at a single meal. Thus,
because they do not need to eat constantly and because by resting
they can conserve energy, carnivores spend a great deal of time each
day sleeping.
The behavior of some animals does appear odd, however. Opossums,
which spend much of their time asleep, may have specialized
in energy conservation as a survival strategy.We humans are average
among species in our sleep time, which is presumably indicative of
an evolutionary pattern in which food gathering was not an overwhelming
preoccupation and predation was not a major concern.
Sleep can contribute to energy conservation in a number of
ways.During sleep, energy is not being expended in moving the body
or supporting posture. The brain is a major user of energy, and so
switching off the brain during sleep, especially NREM sleep, is another
good way to conserve energy. The drop in body temperature
that typically accompanies sleep slows metabolic activity, and so it,
too, contributes to energy conservation.
A good explanation of sleep must explain not only sleep but also
NREM and REM sleep. Before the discovery of REM sleep, Kleitman
suggested that animals have a basic rest–activity cycle (BRAC) that,
for humans, has a period of about 90 min (see Dement, 1972).
0 24
Average time spent in sleep per day (hours)
3 6 9 12 15 18
Opossum
Cat
Rabbit
Rat
Guinea pig
Human
Donkey
21
Figure 12-17
Average Sleep Time Sleep time is
affected both by the amount of time
required to obtain food and by the risk
of predation.
Figure 12-16
Do Not Disturb Biological theories of
sleep suggest that sleep is an energyconserving
strategy and serves other
functions as well, such as staying safe
during the night.
Lynn Hoffman/Photo Researchers
CH12.qxd 2/7/05 2:58 PM Page 465
ls
le
ll
He based his hypothesis on the observation that
human infants have frequent feeding periods between
which they sleep.
As is illustrated in Figure 12-18, the behavior
of adult humans also suggests that activity and
rest are organized into temporal packets. School
classes, work periods, meal times, coffee breaks,
and snack times appear to be divided into intervals
of 90 min or so. The later discovery that REM
sleep occurs at intervals of about 90 min added support to Kleitman’s hypothesis, because
the REM periods could be considered a continuation into sleep of the 90-min
BRAC cycle. The hypothesis now assumes that periods of eating are periods of high
brain activity, just as are periods of REM.
Kleitman proposed that the BRAC rhythm is so fundamental that it cannot be
turned off. Accordingly, in order for a night’s sleep to be uninterrupted by periodic
waking (and perhaps snacking), the body is paralyzed and only the brain is active. To
use an analogy, rather than turning off your car’s engine when you’re stopped at a red
light, you apply the brakes to keep the idling car from moving.
Sleep As a Restorative Process
The idea that sleep has a restorative function is widely held among poets, philosophers,
and the public, as illustrated by Shakespeare in Macbeth’s description of sleep:
Sleep that knits up the ravell’d sleave of care,
The death of each day’s life, sore labour’s bath,
Balm of hurt minds, great nature’s second course,
Chief nourisher in life’s feast.
Sleep as a restorative is also an idea that we can understand from a personal perspective.
Toward the end of the day, we become tired, and, when we awaken from sleep,
we are refreshed. If we do not get enough sleep, we often become irritable. One hypothesis
of how sleep is restorative proposes that chemical events that provide energy
to cells are reduced during waking and are replentished during sleep.
Nevertheless, fatigue and alertness may simply be aspects of the circadian rhythm
and have nothing at all to do with wear and tear on the body or depletion of essential
bodily resources. To evaluate whether sleep is essential for one or another bodily
process, studies of sleep deprivation have been conducted.
Sleep-deprivation studies have not identified any function for which sleep is essential.
One case study on sleep deprivation described by Dement illustrates this point.
In 1965, as part of a science-fair project, a student named Randy Gardner planned to
break the world record of 260 hours (almost 11 days) of consecutive wakefulness with
the help of two classmates, who would keep him awake.
Gardner did break the record and then slept for 14 hours and reported no ill effects.
The world record now stands at a little more than 18 days. A number of reviews
of sleep-deprivation research are consistent in concluding that, at least for these limited
periods of sleep deprivation, no marked physiological alterations ensue.
Although sleep deprivation does not seem to have adverse physiological consequences,
it is associated with poor cognitive performance.Decreased performance contributes
to accidents at work and on the road. The sleep-deprivation deficit does not
manifest itself in an inability to do a task, because sleep-deprived subjects can perform
466 ! CHAPTER 12
Sleep Breakfast
8:00 AM
Coffee
10:00 AM
Sleep–wake
cycle
Lunch
12:00 PM
Snack
3:00 PM
Basic rest–
activity cycle
Dinner
5:00 PM
Snack
7:00 PM
Sleep
Figure 12-18
Behavioral Rhythms Our behavior is
dominated by a basic rest–activity cycle
(red) in which our activity levels change
in the course of the day and by an
NREM–REM sleep cycle (purple) during
the night.
CH12.qxd 2/7/05 2:58 PM Page 466
ls
le
ll
even very complex tasks. Rather, the deficit is revealed when sustained attention is required
and when a task is repetitive or boring.
Even short periods of sleep deprivation, amounting to the loss of a few hours of
sleep, can increase errors on tasks requiring sustained attention. A confounding factor
in cognitive performance is that sleep-deprived subjects will take microsleeps, brief
sleeps lasting a few seconds. During microsleep, subjects may remain sitting or standing,
but their eyelids droop briefly and they become less responsive to external stimuli.
Many people who have driven a car while tired have experienced a microsleep and
awakened just in time to prevent themselves from driving off the road.
Some studies have focused on the selective contributions of REM sleep. To deprive
a subject of REM sleep, researchers allow subjects to sleep but awaken them as they
start to go into REM sleep. REM-sleep deprivation has two effects:
1. Subjects show an increased tendency to go into REM sleep in subsequent sleep sessions,
and so awakenings must become more and more frequent.
2. Subsequent to REM deprivation, subjects experience “REM rebound,” showing
more than the usual amount of REM sleep in the first available sleep session.
Some early reports of REM-deprivation studies stated that subjects could begin to hallucinate
and display other abnormalities in behavior, but these reports have not been
confirmed.
Two kinds of observations, however, suggest no adverse effects of prolonged or
even complete deprivation of REM sleep. Virtually all antidepressant drugs, including
MAO inhibitors, tricyclic antidepressants, and SSRIs, suppress REM sleep either
partly or completely. The clinical effectiveness of these drugs may in fact derive from
their REM-suppressant effects (Vogel et al., 1990). There are no reports of adverse
consequences from prolonged REM deprivation as a consequence of treatment with
antidepressants.
In a number of reported cases, lower-brainstem damage resulted in a complete loss
of REM sleep. Some of these people suffered from locked-in syndrome: they were fully
conscious, alert, and responsive but quadriplegic and mute. Five of seven patients with
locked-in syndrome were repoted to have no REM sleep, without apparent ill effects
(Markand & Dyken, 1976). Patients with more-selective brainstem lesions reportedly
remained ambulatory and verbally communicative, but their REM was abolished. They
lived quite satisfactorily without REM sleep (Osorio & Daroff, 1980).
Sleep and Memory Storage
A fourth explanation of sleep proposes that sleep plays a role in solidifying and organizing
events in memory. One group of experimenters proposes that events are stored
in permanent memory in NREM sleep, whereas another group proposes that REM
sleep fulfills this function.
To examine whether rats dream,Matthew Wilson and Bruce McNaughton (1994)
made use of the finding that many hippocampal cells fire when a rat is in a certain
location in an environment. These place cells are relatively inactive until the rat
passes through a particular place in its environment, then they display a high note of
discharge.
The experimenters trained rats to look for food in a circular container or to search
for food on a four-arm maze.Recordings were made from as many as 100 cells at the same
time in three conditions: during NREM sleep, during a session in the food-searching task,
WHY DO WE SLEEP AND DREAM? ! 467
Microsleep. Brief period of sleep lasting
a second or so.
Locked-in syndrome. Lower brainstem
damage results in a fully conscious, alert,
and responsive condition, but the patient
is quadriplegic and mute.
Place cell. Hippocampal neuron that
fires when a rat is in a certain location in
an environment.
CH12.qxd 2/7/05 2:58 PM Page 467
ls
le
ll
and during NREM sleep after a session in the food-searching task. The experimenters
then used computer methodology to look for cells whose discharge was correlated—that
is, cells that discharged in a consistent temporal relation.
As is illustrated in Figure 12-19, the activity of only a few cells was strongly correlated
in the sleep session that preceded the food-searching task. In the course of food
searching, quite a number of cells discharged at the same time as a rat visited different
locations in the apparatus. In the sleep session that followed the food-searching
experience, correlations that were remarkably similar to those observed in food
searching were observed.
Wilson and McNaughton proposed that, during NREM sleep, the memory of the
previous food-searching experience is stored. Thus, if given another foraging opportunity,
the animal knows where to look for the food.
To determine whether humans’ dreams are related to memory, Pierre Maquet
and his coworkers in Belgium trained subjects on a serial reaction task and observed
regional blood flow in the brain with PET scans during training and during REM
sleep on the subsequent night (Maquet et al., 2000). The subjects faced a computer
screen on which six positional markers were displayed. The subjects were to push one
of six keys when a corresponding positional marker was illuminated. The subjects did
not know that the sequence in which the positional markers were illuminated was
predetermined.
Consequently, as training progressed, the subjects indicated that they were
learning because their reaction time improved on trials on which one positional
marker was correlated with a preceding marker. On the PET-scan measures of brain
activation, a similar pattern of neocortical activation was found during task acquisition
and during REM sleep (Figure 12-20). On the basis of this result, Maquet and
coworkers suggest, first, that the subjects were dreaming about their learning experience
and, second, that the replay during REM strengthened the memory of the
task.
468 ! CHAPTER 12
NREM sleep
before food
search
Search
for food
NREM sleep after
food search
Dots represent
individual
hippocampal cells.
Red lines connecting dots represent cells that
fire at the same time. The thicker the line,
the higher the correlation of cell activity.
The correlation of cell
activity during sleep is
similar to that during
searches for food when
previously awake.
Figure 12-19
Neural Replay? The activity of
hippocampal cells suggests that rats
dream about previous experiences. The
dots on the periphery of the circles
represent the activity of 42 hippocampal
cells recorded at the same time during
(1) slow-wave sleep before a foodsearching
task, (2) the food-searching
task, and (3) slow-wave sleep after the
task. No strong correlations between
cells emerged during the slow-wave
sleep that preceded the food-searching
task, but correlations between cells
during the food search and during the
subsequent slow-wave sleep were
strong. Adapted from “Reactivation of
Hippocampal Ensemble Memories During Sleep,”
by M. A. Wilson and B. L. McNaughton, 1994,
Science, 165, p. 678.
(A) Reaction-time task (B) REM sleep that night
Subjects are
trained on a
reaction-time
task, and brain
activity is
recorded with
PET.
Subjects display
a similar
pattern of
brain activity
during
subsequent
REM sleep.
Figure 12-20
Do We Store Memories
During REM Sleep? Adapted
from “Experience Dependent
Changes in Cerebral Activation
During Human REM Sleep,” by P.
Maquet et al., 1998, Nature
Neuroscience, 3, p. 832.
CH12.qxd 2/7/05 2:58 PM Page 468
ls
le
ll
The results of some natural experiments raise questions about the memory-storing
functions of sleep. There are many documented cases of people who sleep less than
a couple of hours each day yet remain active and healthy, presumably with normal
memory. These cases raise the question of why most people need much longer periods
of sleep.
Consider the case reported of a 33-year-old man who suffered a head injury at age
20 and subsequently displayed little REM sleep, as documented in sleep-recording sessions
in a sleep laboratory (Lavie et al., 1984). The lack of REM sleep did not appear to
cause serious effects: the subject completed high school, attended law school, and subsequently
practiced law.
Human infants spend a great deal of time sleeping in their first year or so but remember
nothing of this period of life, although they are amassing motor skills. And
the sloth is one of nature’s great sleepers but is not noted among its great learners.
Perhaps sleep facilitates plastic changes in the brain more generally and facilitates
memory because memory depends on brain plasticity. Nevertheless, memory-storing
explanations of sleep are extremely interesting and are being hotly debated (e.g., see
Vertes, 2004, versus Walker & Stickgold, 2004). Sleep may also facilitate the normal
function of the immune system, which also undergoes changes in its function in defense
against disease.
NEURAL BASIS OF SLEEP
The idea that the brain contains a sleep-inducing substance has long been popular and
is reinforced by knowledge that a variety of chemical agents induce sleep. Such substances
include sedative-hypnotics and morphine (Chapter 7).A twist on this idea held
that the body secretes a chemical that induces sleep and that can be removed only by
sleeping.
The hormone melatonin, secreted from the pineal gland during the dark phase of
the light–dark cycle, causes sleepiness and is taken as an aid for sleep, and so it might be
thought to be the sleep-producing substance. Sleep, however, survives the removal of
the pineal gland. Thus,melatonin, and many other chemical substances,may only contribute
to sleep, not cause it. If any chemical actually regulates sleep, it has not yet been
identified (see ”Synchronizing Biorhythms at the Molecular Level” on page 452).
In fact, experimenters have obtained evidence that sleep is not produced by a compound
circulating in the bloodstream.When dolphins and birds sleep, only one hemisphere
sleeps at a time. This ability presumably allows an animal’s other hemisphere to
remain behaviorally alert. This observation also strongly suggests that sleep is produced
by the action of some region within each hemisphere.
In this section, we consider two points about the neural basis of sleep. First, we
examine evidence that sleep is produced by the activity of a slave oscillator of the
In Review .
Among the explanations put forward concerning why we sleep, the biological explanation—
that sleep is an adaptive strategy for conserving energy during times when food resources
are hard to obtain—has replaced the passive explanation—that sleep results from
lack of sensory stimulation. Scientists are examining the ideas that sleep is a restorative
process and that sleep has a role in storing and sorting memory, but so far the evidence
in favor of these ideas is not conclusive.
WHY DO WE SLEEP AND DREAM? ! 469
Sloth
CH12.qxd 2/7/05 2:58 PM Page 469
ls
le
ll
suprachiasmatic nucleus (see Figure 12-8). Second, we look at evidence that the various
events associated with sleep, including events associated with REM and NREM
sleep, are controlled by a number of different brainstem nuclei.
Reticular Activating System and Sleep
A dramatic experiment and a clever hypothesis by Giuseppe Moruzzi and Horace
Magoun (1949) provide the beginnings of an answer to the question of which areas of
the brain regulate sleep. Moruzzi and Magoun were recording the cortical EEG from
anesthetized cats while electrically stimulating the cats’ brainstems. They discovered
that, in response to the electrical stimulation, the large, slow cortical EEG typical of the
condition of anesthesia was dramatically replaced by the low-voltage, fast-wave EEG
typical of waking.
The waking pattern of EEG activity outlasted the period of stimulation, demonstrating
that the pattern was produced by the activity of neurons in the region of the
stimulating electrode. During the “waking period,” the cat did not become behaviorally
aroused, because it was anesthetized, but its cortical EEG appeared to indicate
that it was awake. This EEG pattern is referred to as a desynchronized EEG, meaning
that the large, synchronized waves of sleep are replaced by low-voltage, fast activity
brain waves of waking.
Findings from subsequent experiments by Moruzzi and Magoun and by others
showed that a desynchronized EEG could be induced from a large area running
through the center of the brainstem. Anatomically, this area is composed of a mixture
of cell nuclei and nerve fibers that form a reticulum (from the Latin word rete, meaning
“net”; described in Chapter 2). Moruzzi and Magoun named this brainstem area
the reticular activating system (RAS) and proposed that it is responsible for sleep–
waking behavior. The location of the RAS is illustrated in Figure 12-21.
We know that, if someone disturbs us when we are asleep, we usually wake up. To
explain how sensory stimulation and the RAS are related, Moruzzi and Magoun proposed
that sensory pathways entering the brainstem have collateral axons that synapse
with neurons in the RAS. They proposed that sensory stimulation is conveyed to RAS
neurons by these collaterals, and then RAS neurons produce the desynchronized EEG
by axons that project to the cortex.
The results of subsequent experiments with sleeping cats show that electrical
stimulation produces waking EEG activity and behavioral arousal in animals, just as
if sensory stimulation had been used to wake them up. Moruzzi and Magoun further
proposed that the cortex sends axons to the RAS, providing a route for people to stimulate
their own reticular activating systems to stay awake. In sum, both sensory stimulation
and conscious effort could activate the RAS to maintain waking.
Two other lines of experimental evidence support the idea that the RAS is responsible
for desynchronized EEGs. Because Moruzzi and Magoun could possibly have
stimulated various sensory pathways passing through the brainstem, it was necessary
to demonstrate that brainstem neurons and not sensory-pathway stimulation produced
the waking EEG. When experimenters cut the brainstem just behind the RAS,
thereby severing incoming sensory pathways, RAS stimulation still produced a desynchronized
EEG.
This result strengthened the argument that RAS neurons, not sensory pathways
running through the region, are responsible for producing a desynchronized EEG. Furthermore,
if the cut was made through the brainstem just in front of the RAS, the desynchronized
EEG was no longer obtained in response to electrical stimulation of the
470 ! CHAPTER 12
Reticular activating system (RAS).
Large reticulum (mixture of cell nuclei
and nerve fibers) that runs through the
center of the brainstem.
Coma. Prolonged state of deep
unconsciousness resembling sleep.
Reticular activating
system (RAS)
Hypothalamus
Cortex
Sensory
information
Figure 12-21
Sleep–Wake Controller The reticular
activating system, a region in the middle
of the brainstem, is characterized by a
mixture of cell bodies and fiber
pathways. Stimulation of the RAS
produces a waking EEG, whereas
damage to it produces a slow-wave,
sleeplike EEG.
CH12.qxd 2/7/05 2:58 PM Page 470
ls
le
ll
RAS. Together these experiments demonstrate that RAS neurons acting through axons
projecting to the cortex produce the waking EEG.
A different line of evidence obtained from humans who have suffered brainstem
injury supports this conclusion. Damage that affects the RAS results in coma, a state
of deep unconsciousness resembling sleep. In a well-publicized case, after taking a
minor tranquilizer and having a few drinks at a birthday party, a 21-year-old woman
named Karen Ann Quinlan sustained RAS damage that put her in a coma (Quinlan &
Quinlan, 1977). She was hospitalized, placed on a respirator to support breathing, and
fed by tubes. Her family fought a protracted legal battle to have her removed from life
support, which was finally won before the Supreme Court of New Jersey. Even after
having been removed from life support, however, Quinlan lived for 10 more years in a
perpetual coma.
Despite substantial evidence that the RAS has a role in sleep–waking behavior, attempts
to localize sleep to a particular structure or group of neurons within the RAS
have not been successful. Findings from many studies demonstrate that discrete lesions
at various locations within the RAS can produce periods of sleep that last for days; but,
with care, animals in the laboratory and human brain-injured patients recover from
these acute symptoms. These findings suggest that sleep–waking behavior is due to the
activity of a diffuse network of fibers and cells rather than to regulation by a single nucleus.
As described in the next section, patterns of a waking EEG are produced through
at least two routes.
Neural Basis of EEG Changes Associated
with Waking
A series of experiments performed on rats by Case Vanderwolf and his coworkers
(Vanderwolf, 1988) suggested that nuclei in the midbrain and forebrain are responsible
for producing the waking EEG pattern of the neocortex. Figure 12-22 illustrates the
location of these structures. Both send neural pathways into the neocortex, where they
make diffuse connections with cortical neurons.
The basal forebrain contains large cholinergic cells. These neurons secrete acetylcholine
(ACh) from their terminals onto neocortical neurons to stimulate a waking
EEG (beta rhythm). The midbrain structure, the median raphé, contains serotonin
(5-HT) neurons whose axons also project diffusely to the neocortex, where they also
stimulate neocortical cells to produce a beta rhythm, recorded as a waking EEG.
Although both pathways produce a very similar pattern of waking EEG activity, the
relations of the two types of desynchronized EEG to behavior are different. If the
activity of the cholinergic projection is blocked by drugs or by lesions to the cells of the
basal forebrain, the waking EEG that is normally recorded from an immobile rat is
replaced by EEG activity resembling that of NREM sleep. If the rat
walks or is otherwise active, a waking EEG is obtained from the neocortex.
These findings suggest that the cholinergic EEG is responsible
for the waking EEG when the rat is still and alert, whereas the serotonergic
activation is additionally responsible for the waking EEG
when the animal moves.
It is important to note that neither the basal forebrain system nor
the median raphé system is responsible for behavior. In fact, if both
structures are pharmacologically or surgically destroyed, a rat can still
stand and walk around. Its neocortical EEG, however, resembles that of
a sleeping animal.
WHY DO WE SLEEP AND DREAM? ! 471
Rest Alert Rest Move
Basal
forebrain (ACh)
Median
raphé (5-HT)
Figure 12-22
Brain Activators In the rat, basal
forebrain ACh neurons produce an
activated EEG pattern when a rat is alert
but immobile. The 5-HT raphé neurons
of the midbrain produce an activated
EEG pattern when the rat moves.
CH12.qxd 2/7/05 2:58 PM Page 471
ls
le
ll
As long as one of the activating systems is producing a waking EEG, rats can learn
simple tasks. If both systems are destroyed, however, an animal, although still able to
walk around, is no longer able to learn or display intelligent behavior. In a sense, the
cortex is like a house in which the lights are powered by two separate power sources:
both must fail for the house to be left in darkness, but, if at least one source is operating,
the lights will be on.
We do not know if the basal forebrain and median raphé produce the same two
desynchronized EEG patterns in humans as they do in rats, but they likely do. Consequently,
when we are alert, the cholinergic neurons are likely to be active and, when we
move, the serotonin neurons are likely to be additionally active. You may have had the
experience, when you felt sleepy in a class or behind the wheel of a car, of being able
to wake yourself up by moving—shaking your head or stretching. Presumably, your
arousal level decreased as your cholinergic neurons became inactive; but, when you
moved, your serotonergic neurons became active and restored your level of arousal.
When we enter sleep, both cholinergic and serotonergic neurons become less active, allowing
slow waves to emanate in the cortex.
Neural Basis of REM Sleep
When we were looking at evidence related to the function of REM
sleep, we considered a number of clinical cases in which people who
had suffered brainstem damage no longer displayed REM sleep.
This observation suggests that REM sleep is produced by the action
of a neural area distinct from the RAS, which produces NREM
sleep.
Barbara Jones (1993) and her colleagues described a group of
cholinergic neurons known as the peribrachial area, which appears
to be implicated in REM sleep. This area is located in the dorsal part of the brainstem
just anterior to the cerebellum (Figure 12-23).
Jones selectively destroyed these cells by spraying them with neurotoxin kainic
acid and found that REM sleep in her experimental animal subjects was drastically
reduced. This result suggests that the peribrachial area is responsible for producing
REM sleep and REM-related behaviors. The peribrachial area extends into a more
ventrally located nucleus called the medial pontine reticular formation (MPRF).
Lesions of the MPRF also abolish REM sleep, and injections of cholinergic agonists
(drugs that act like ACh) into the MPRF induce REM sleep. Thus, both the peribrachial
area and the MPRF, illustrated in Figure 12-23, take part in the production of REM
sleep.
If these two brain areas are responsible for producing REM sleep, how do other
events related to REM sleep take place? Such events include:
EEG pattern similar to waking EEG;
rapid eye movements, or REM;
sharp EEG spikes that are recorded from the pons, the lateral geniculate nucleus,
and the visual cortex and are called PGO (pons, geniculate, occipital) waves after the
structures in which they are found; and
atonia, or absence of muscle tone due to the inhibition of motor neurons.
These REM-related activities are likely to be found in humans, although, at present,
PGO waves have not been confirmed in humans.
472 ! CHAPTER 12
Peribrachial area. Cholinergic nucleus
in the dorsal brainstem having a role in
REM sleep behaviors; projects to medial
pontine reticulum.
Medial pontine reticular formation
(MPRF). Nucleus in the pons
participating in REM sleep.
Barbara Jones
Peribrachial
area
Medial pontine
reticular formation
Figure 12-23
Brainstem Nuclei Responsible for
REM Sleep Damage to either the
peribrachial area or the medial pontine
formation will reduce or abolish REM
sleep.
CH12.qxd 2/7/05 2:58 PM Page 472
ls
le
ll
One explanation of how other REM-related activities take place is illustrated
in Figure 12-24. The MPRF sends projections to basal forebrain cholinergic neurons
to activate them, resulting in the production of the activated EEG of the cortex.
The MPRF area also excites the PGO pathway to produce PGO waves and eye
movements. Finally, the atonia of REM sleep is produced by the MPRF through
a pathway that sends input to the subcoerulear nucleus, located just behind it. The
subcoerulear nucleus excites the magnocellular nucleus of the medulla, which
sends projections to the spinal motor neurons to inhibit them so that paralysis is
achieved during the REM-sleep period.
French researcher Michael Jouvet (1972) observed that cats with lesions in
the subcoerulear nucleus display a remarkable behavior when they enter REM
sleep. Rather than stretching out in the atonia that typically accompanies REM
sleep, they stood up, looked around, and made movements of catching an imaginary
mouse or running from an imaginary threat. If cats dream about catching
mice or dream about escaping from a threat, then these cats appear to have been
acting out their dreams.
SLEEP DISORDERS
Occasional disturbances of sleep are annoying and may result in impaired performance
during the following day. About 15 percent of people complain of ongoing sleep problems;
an additional 20 percent complain of occasional sleep problems. As people age,
the incidence of complaints about sleep problems increases. In this section, we consider
abnormalities of NREM sleep and REM sleep.
Disorders of Non-REM Sleep
The two most common sleep disorders are insomnia, prolonged inability to sleep, and
narcolepsy (from the Greek narco, “a stupor,” and lepsy, “to be seized”), uncontrollably
falling asleep at inconvenient times. Both are considered disorders of slow-wave sleep.
Insomnia and narcolepsy are related, as anyone who has stayed up late at night can confirm:
a short night’s sleep is often accompanied by a tendency to fall asleep at inconvenient
times the next day.
Our understanding of insomnia is complicated by a large variation in how much
time people spend asleep. Some short sleepers may think that they should sleep more,
and some long sleepers may think that they should sleep less; yet, for each, the sleeping
pattern may be appropriate. It is also possible that, for some people, circadian
rhythms are disrupted by subtle life-style choices. Staying up late, for example,may set
In Review .
Separate neural regions are responsible for sleep. The reticular activating system (RAS) in
the central region of the brainstem is responsible for NREM sleep, whereas the peribrachial
area and the medial pontine reticular formation (MPRF) are responsible for REM sleep. The
last two areas, through activating pathways to the neocortex and spinal cord, are responsible
for producing the waking EEG and the muscular paralysis that are associated with
REM sleep.
WHY DO WE SLEEP AND DREAM? ! 473
Peribrachial area
initiates REM
Loss of muscle tone
produced by the
subcoerulear nucleus
exciting the magnocellular
nucleus of the medulla
Spinal motor
neurons inhibited
Desynchronized EEG
in neocortex produced
by basal forebrain
PGO
waves
Medial pontine reticular
formation produces REMrelated
activities
Figure 12-24
Neural Control of REM Sleep
Insomnia. Disorder of slow-wave sleep
resulting in prolonged inability to sleep.
Narcolepsy. Slow-wave sleep disorder
in which a person uncontrollably falls
asleep at inappropriate times.
CH12.qxd 2/7/05 2:58 PM Page 473
ls
le
ll
a person’s circadian rhythm forward, encouraging a cascade of late sleep followed by
still later staying up.
Some sleep problems are brought on by shift work or by jet lag, situations described
earlier in the chapter. Other common causes of sleep disorders are stress, long
hours of work, and irregular life styles. Just worrying about insomnia is estimated to
play a major role in 15 percent of insomnia cases.
Sleep disorders are a complicating factor in other conditions, including depression.
People who are depressed may sleep too much or too little. Anxiety and depression
may account for about 35 percent of insomnias. There are also quantitative
differences in the sleep of depressed patients, because they enter REM sleep very
quickly. Entering REM sleep quickly, however,may be secondarily related to sleep deprivation,
rather than being related directly to depression, because people who are sleep
deprived also enter REM very quickly. Irregular sleeping patterns are also common in
schizophrenia.
Insomnia may be brought on by sedative-hypnotic drugs, including seconal, sodium
amytal, and many minor tranquilizers. These “sleeping pills” do help people get
to sleep, but they cause additional problems. People may sleep under one of these
drugs, but they are likely to feel groggy and tired the next day, which defeats the purpose
of taking the drug. In addition, people develop tolerance to these medications, become
dependent on them, and display rebound insomnia when they stop taking them.
A person may increase the dose of the drug each time the drug fails to produce the desired
effect. The syndrome in which patients unsuccessfully attempt to sleep by increasing
their dosage of a drug is called drug-dependency insomnia.
Like many other people, you may have had the experience of being suddenly overcome
by an urge to sleep at an inconvenient time, such as while attending a lecture. For
some people, such experiences with narcolepsy are common and disruptive. J. S., a junior
in college, sat in the front row of the classroom for his course on the brain; within
a few minutes of the beginning of each class, he dropped off to sleep. The instructor
became concerned about his abilities as a lecturer, but one day he heard another instructor
describe the sleeping behavior of a student who turned out to be J. S. The instructor
then asked J. S. to stay after class to discuss his sleeping behavior.
J. S. reported that sleeping in classes was a chronic problem. Not only did he sleep
in class, but he fell asleep whenever he tried to study. He even fell asleep at the dinner
table and in other inappropriate locations. His sleeping problem had made it a challenge
to get through high school and was making it very difficult for him to pass his
college courses.
About 1 percent of people suffer from narcolepsy, which takes a surprising number
of forms. One cause of narcolepsy maybe be mutations in a gene that produces
hypocretin/orexin peptides. Mutations in this gene or in the gene that makes the receptors
for these compounds have been related to narcolepsy (Mignot, 2004).
J. S. had a form of narcolepsy in which he fell asleep while sitting still, and his sleeping
bouts consisted of brief bouts of NREM sleep lasting from 5 to 10 min. This pattern
was very similar to napping and to dropping off to sleep in class after a late night,
but it was distinguishable as narcolepsy by its frequency and by the disruptive effect
that it had on his academic career. J. S. eventually discussed his problem with his physician
and received a prescription for Ritalin, an amphetamine-like drug that stimulates
dopamine transmission (Chapter 7). The treatment proved very helpful.
Some people who suffer from daytime sleepiness attend sleep clinics to get help in
sleeping better at night. Studies of narcoleptic people in sleep clinics have resulted in
one surprising discovery concerning one of the causes of narcolepsy. “Sleep Apnea” describes
a person who had to wake up to breathe; his sleep apnea left him extremely
tired and caused him to nod off in the daytime.
474 ! CHAPTER 12
Drug-dependency insomnia.
Condition resulting from continuous use
of “sleeping pills”; drug tolerance also
results in deprivation of either REM or
NREM sleep, leading the user to increase
the drug dosage.
Sleep apnea. Inability to breathe during
sleep; person has to wake up to breathe.
Go to the Web site at www.worth
publishers.com/kolb/chapter12 to
learn more about narcolepsy.
CH12.qxd 2/7/05 2:58 PM Page 474
ls
le
ll
Disorders of REM Sleep
Recall from the description of REM sleep that it is associated with muscular atonia and
dreaming. REM-sleep atonia can occur when a person is not asleep, which happened
to L. M., a college senior who, after hearing a lecture on narcoleptic disorders, recounted
the following experience.
WHY DO WE SLEEP AND DREAM? ! 475
Sleep Apnea
Focus on Disorders
The first time I went to a doctor for my insomnia, I
was twenty-five—that was about thirty years ago.
I explained to the doctor that I couldn’t sleep; I had
trouble falling asleep, I woke up many, many times
during the night, and I was tired and sleepy all
day long. As I explained my problem to him, he
smiled and nodded. Inwardly, this attitude infuriated
me—he couldn’t possibly understand what I
was going through. He asked me one or two questions:
Had any close friend or relative died recently?
Was I having any trouble in my job or at
home? When I answered no, he shrugged his shoulders
and reached for his prescription pad. Since
that first occasion I have seen I don’t know how
many doctors, but none could help me. I’ve been
given hundreds of different pills—to put me to sleep
at night, to keep me awake in the daytime, to calm
me down, to pep me up—have even been psychoanalyzed.
But still I cannot sleep at night. (In Dement,
1972, p. 73).
This patient went to the Stanford University Sleep Disorders
Clinic in 1972. Recording electrodes were attached to him
so that brain, muscle, eye, and breathing activity could be
recorded while he slept (see Figure 12-11). The experimenters
were amazed to find that he had to wake up to
breathe.
They observed that he would go for more than a minute
and a half without breathing, wake up and gasp for breath,
and then return to sleep, at which time the sequence was repeated.
Such sleep apnea (a, “not,” and pnea, “breathing”)
may be produced by a central problem, such as a weak command
to the respiratory muscles, or it may be obstructive,
caused by collapse of the upper airway. When people suffering
from sleep apnea stop breathing, they either wake up
completely and have difficulty getting back to sleep or they
have repeated partial awakenings throughout the night to
gasp for breath.
Because sufferers are apparently unaware of their sleep
apnea, it must be diagnosed by someone who watches them
sleep. Sleep apnea affects all ages and both sexes, and 30 percent
of people older than 65 years of age may have some form
of it. Sleep apnea can even occur in children, in whom it has
been associated with daytime behavioral disorder (Rosen et
al., 2004). Sleep apnea is thought to be more common among
people who are overweight and who snore, conditions in
which air flow is restricted.
Treatments for sleep apnea include surgery or appliances
that expand the upper airway, weight loss, and face
masks that deliver negative pressure to open the airway.
Sleep apnea may also be related to sudden infant death syndrome
(SIDS), or crib death, in which otherwise healthy infants
inexplicably die in their sleep.
Breathing rate and blood-oxygen level recorded from a person with
sleep apnea during REM sleep. Blood oxygen increased after each
breath but then continued to fall until another breath was taken.
This person inhaled only 4 times in the 6-min period; a normal
sleeper would breathe more than 60 times in the same interval.
Time (min)
0 1 2 3 4 5 6
Breathing
Four breaths in
6 minutes
Blood
oxygen
CH12.qxd 2/7/05 2:58 PM Page 475
ls
le
ll
476 ! CHAPTER 12
She had just gone to sleep when her roommate came into their room. She woke
up and intended to ask her roommate if she wanted to go skating the next morning
but found herself completely unable to speak. She tried to turn her head to follow her
roommate’s movements across the room but found that she was paralyzed. She had
the terrifying feeling that some kind of monstrous creature was hiding in the bathroom
waiting for her roommate. She tried to cry out but could produce only harsh,
gurgling noises. Her roommate knocked her out of her paralysis by hitting her with a
pillow.
This form of narcolepsy, called sleep paralysis, is extremely common. In informal
class surveys, almost a third of students report that they have had such an experience.
The atonia is typically accompanied by a feeling of dread or fear. It seems likely that,
in sleep paralysis, a person has entered REM sleep and atonia has occurred, but
the person remains partly conscious or has partly awakened.
The atonia of REM sleep may also occur while a person is awake; this
form is called cataplexy (from the Greek cata, “to fall,” and plexus, “seized”).
In cataplexy, an awake, alert person suddenly falls to the floor, atonic.
Cataplexy is frequently reported to be triggered by excitement or laughing.
Suddenly, the jaw drops, the head sinks, the arms go limp, the legs
buckle, and the person falls down. The collapse can be so sudden that
there is a real risk of injury.
While in an atonic condition, the person may see imaginary
creatures or hear imaginary voices, called hypnogogic (Greek
hypno, “sleep,” and agogic, “leading into”) hallucinations. People
who fall into a state of cataplexy with hypnogogic hallucinations
give every appearance of having fallen into REM sleep
while remaining conscious.
Conditions in which REM-sleep atonia occurs frequently
may have a genetic basis. In 1970, William Dement
was given a litter of Doberman pinscher dogs and, later, a
litter of Labrador retrievers, all of whom had cataplexy. The
disease is transmitted as a recessive trait; so, to develop it, a
dog must inherit the gene from both its mother and its father.
The descendants of those dogs provide animal models
for investigating the neural basis of the disease as well as its
treatment.
When a dog with cataplexy is excited, such as when it is
running for a piece of food, it may suddenly collapse, as is
illustrated on the left in Figure 12-25. Jerome Siegel (2004)
investigated the cause of narcolepsy in dogs and found that
Figure 12-25
Cataplexy In both dog and human, an attack of
catalepsy causes the head to droop and the back and
legs to sag and can progress to a complete loss of muscle
tone while the person or dog is awake and conscious.
Cataplexy is distinct from the sleep attacks that people
hear and remember what is said around them, and
dogs can track a moving object with their eyes. James
Aronovsky (dog sequence), Joel Deutsch (human sequence),
Slim Films.
CH12.qxd 2/7/05 2:58 PM Page 476
ls
le
ll
neurons in the subcoerulear nucleus become inactive and neurons in the magnocellular
nucleus of the medulla become active during attacks of cataplexy, just as they do
during REM sleep. For some reason, the neurons responsible for paralysis during
REM were producing paralysis during waking.
On the basis of anatomical examinations of the brains of narcoleptic dogs, Siegel
suggested that the death of neurons in the amygdala and adjacent forebrain areas
is a one-time event just before the onset of the disease early in life. Presumably,
the loss of these neurons somehow results in the loss of inhibition in the brainstem
areas that produce paralysis. It is important to note that, although a genetic basis
for cataplexy has been identified in dogs, there is as yet no evidence that all cases of
human cataplexy are genetic. Like some forms of narcolepsy, cataplexy is treatable
with Ritalin.
Recall Jouvet’s experiment, in which he reported that cats with lesions to the subcoerulear
region of the brainstem entered REM sleep without the accompanying atonia
and apparently acted out their dreams. A similar condition has been reported in people
and may either have a genetic basis or be caused by brain damage. The condition has
been named REM without atonia. The behavior of people who have REM without atonia
suggests that they are acting out their dreams. The following two accounts are those
of a 67-year-old patient (Schenck et al.,1986):
I was on a motorcycle going down the highway when another motorcyclist
comes up alongside me and tries to ram me with his motorcycle. Well, I decided,
I’m going to kick his motorcycle away and at that point my wife woke
me up and said, “What in heavens are you doing to me?” because I was kicking
the hell out of her.
I had a dream where someone was shooting at me with a rifle and it was in a
field that had ridges in it, so I decided to crawl behind a ridge—and I then
had a gun too—and I look over the ridge so when he showed up I would
shoot back at him and when I came to [i.e., awakened] I was kneeling alongside
the bed with my arms extended like I was holding the rifle up and ready
to shoot.
In both dreams the patient had vivid pictorial images, but he heard nothing and
he felt afraid. Although a large number of patients who have had such experiences
have been described, most are elderly and suffer from brain injury or other brainrelated
disorders. REM without atonia can be treated with benzodiazepines, which
block REM sleep.
Sleep and Consciousness
Many scientists interested in the neural basis of consciousness study sleep and sleeprelated
disorders. Clearly, the many different qualities and stages of waking and sleeping
suggest that consciousness is not a unitary condition, either neurally or behaviorally.
Rather, we experience a variety of “states of consciousness,” some of which can occur
simultaneously.
René Descartes, whose theory of the duality of body and mind is described in
Chapter 1, conceived of his idea of a mind through a dream. He dreamed that he was
interpreting the dream as it occurred. Later, when awake, he reasoned that, if he could
think and analyze a dream while asleep, the mind must be able to function during both
waking and sleeping. Therefore it must be independent of the body that undergoes
sleeping and waking transitions.
WHY DO WE SLEEP AND DREAM? ! 477
Sleep paralysis. Inability to move
during deep sleep owing to the brain’s
inhibition of motor neurons.
Cataplexy. Form of narcolepsy linked to
strong emotional stimulation in which an
animal loses all muscle activity or tone,
as if in REM sleep, while awake.
Hypnogogic hallucination. Dreamlike
event at the beginning of sleep or while a
person is in a state of cataplexy.
CH12.qxd 2/7/05 2:58 PM Page 477
ls
le
ll
More-recent research sheds additional light on consciousness. For example, what
we colloquially refer to as waking comprises at least two different states: alert consciousness
and consciousness with movement. People attempting to go to sleep or attempting
to stay awake appear to realize that they can take advantage of these different
conditions to achieve their objective.
People who are tired and wish to fall asleep usually seek out a dark, quiet room,
where they lie still. In doing so, they are removing themselves from a condition of
“moving consciousness.”People who want to stay awake, especially if they are tired, can
apparently do so as long as they keep moving. By walking around and otherwise remaining
active, they can stay awake indefinitely.
Similarly, sleep consists of a number of NREM conditions and a REM condition.
People in both NREM sleep and REM sleep are at least in some sense conscious
when they dream. Dream consciousness can also occur in conjunction with waking
consciousness, as witnessed by reports that people who fall into a state of cataplexy
are conscious of being awake while experiencing the visual and emotional features of
dreams when they have hypnogogic hallucinations.
Allan Hobson reported the peculiar symptoms that he suffered after a brainstem
stroke (Hobson, 2002). For the first 10 days after the lesion he suffered from complete
insomnia and experienced neither REM nor non-REM sleep. He did experience
sudden visual hallucinations that had a dreamlike quality whenever he closed
his eyes, however. This experience suggested that eye closure is sufficient to produce
the visual components of REM sleep but with neither loss of consciousness nor
paralysis. He eventually recovered normal sleeping patterns and the hallucinations
stopped.
Besides being a source of insight into the neural basis of consciousness, the study
of sleep states and events may help to explain some psychiatric and drug-induced
conditions. For example, among the symptoms of schizophrenia are visual and auditory
hallucinations. Are these hallucinations dream events that occur unexpectedly
during waking? Many people who take hallucinogenic drugs such as LSD report
that they have visual hallucinations. Does the drug initiate the visual features of
dreams? People who have panic attacks suffer from very real fright that has no obvious
cause. Are they experiencing the fear attacks that commonly occur during sleep
paralysis and cataplexy? The answers to these questions are incomplete, but the similarities
in symptoms between some waking and some sleeping conditions do suggest
that some waking disturbances may be sleep events that occur when a person is
awake.
In Review .
Disorders of NREM sleep include insomnia, in which a person has difficulty falling
asleep at night, and narcolepsy, in which a person falls asleep involuntarily in the
daytime. Treating insomnia with sleeping pills, usually sedative hypnotics, may cause
drug-dependent insomnia, in which progressively higher doses must be taken to
achieve sleep. Disorders of REM sleep include sleep paralysis, in which a person
awakes but is paralyzed and experiences a sense of fear, and cataplexy, in which a person
may lose muscle tone and collapse in the daytime. Cataplexy may be associated
with hypnogogic hallucinations, in which a person experiences dreams while paralyzed
but awake. The study of sleep suggests that consciousness is not unitary but can take
various forms.
478 ! CHAPTER 12
CH12.qxd 2/7/05 2:58 PM Page 478
ls
le
ll
SUMMARY
What are biological rhythms? Biorhythms are cyclic behavior patterns of varying
length displayed by animals, plants, and even single-celled organisms. Mammals display
a number of biological rhythms including circadian, or daily, rhythms and circannual,
or yearly, rhythms. In the absence of environmental cues, circadian rhythms
are free running, lasting a little more or a little less than their usual period of 24 hours,
depending on the individual organism or the environmental conditions. Cues that
reset a biological clock to a 24-hour rhythm are called Zeitgebers.
What is a biological clock? A biological clock is a neural structure responsible for producing
rhythmic behavior. Among the biological clocks in the brain are the suprachiasmatic
nucleus and the pineal gland. The suprachiasmatic nucleus is the mammalian
biological clock responsible for circadian rhythms, and it has its own free-running
rhythm with a period that is a little more or a little less than 24 hours. Stimuli from the
environment such as sunrise and sunset entrain the free-running rhythm so that its period
is 24 hours.
How does a biological clock keep time? Neurons of the suprachiasmatic nucleus are
active in the daytime and inactive at night. These neurons display their rhythmicity
when disconnected from other brain structures,when removed from the brain and cultured
in a dish, and after having been cultured in a dish for a number of generations.
When reimplanted into a brain without a suprachiasmatic nucleus, they restore the animal’s
circadian rhythms. The different aspects of circadian rhythms of neurons, including
their period, are under genetic control.
How is sleep measured? Sleep events are measured by recording the brain’s activity
to produce an electroencephalogram, or EEG; muscular activity to produce an electromyogram,
or EMG; and eye movements to produce an electrooculogram, or EOG.
A typical night’s sleep consists of four stages, as indicated by physiological measures,
which take place in a number of cycles in the course of the night. During REM
sleep, the EEG has a waking pattern, and the sleeper displays rapid eye movements.
Stages of sleep in which the EEG has a slower rhythm are called non-REM (NREM)
sleep. Intervals of NREM sleep and REM sleep alternate four or five times each night;
the duration of NREM sleep is longer in the early part of sleep, whereas the duration
of REM sleep is longer in the later part of sleep period. These intervals also vary
with age.
What events are associated with REM sleep? A sleeper in slow-wave sleep has muscle
tone, may toss and turn, and has dreams that are not especially vivid. A sleeper
in REM sleep has no muscle tone and so is paralyzed and has vivid dreams whose
duration coincides with the duration of the REM period. The activation–synthesis
hypothesis proposes that dreams are not meaningful and are only a by-product of
the brain’s state of excitation during REM. The evolutionary hypothesis suggests
that dreams evolved as a mechanism to cope with real threats and fears posed by the
environment.
Why do we sleep? Several theories of sleep have been advanced, including the propositions
that sleep results from the absence of sensory stimulation, that it is a biological
adaptation that conserves energy resources, and that it is a restorative process that
fixes wear and tear in the brain and body. Sleep may also organize and store each day’s
memories.
What is the neural basis of sleep? Separate neural regions of the brain are responsible
for NREM and REM sleep. The reticular activating system (RAS) located in the central
WHY DO WE SLEEP AND DREAM? ! 479
CH12.qxd 2/7/05 2:58 PM Page 479
ls
le
ll
area of the brainstem is responsible for NREM sleep. If it is stimulated, a sleeper
awakes; if it is damaged, a person may enter a condition of coma. The peribrachial area
and the medial pontine reticular formation of the brainstem are responsible for REM
sleep. If these areas are damaged, REM sleep may no longer occur. Pathways from these
areas project to the cortex to produce the cortical activation of REM and to the brainstem
to produce the muscular paralysis of REM.
What disorders are associated with sleep? Disorders of NREM sleep include insomnia,
the inability to sleep at night, and narcolepsy, inconveniently falling asleep in the
daytime. The administration of sedative hypnotics to induce sleep may induce drugdependency
insomnia, a sleep disorder in which progressively larger doses of the drug
are required to produce sleep. Disorders of REM sleep include sleep paralysis, in which
a dreaming person awakens but remains unable to move and sometimes feels fear and
dread. Cataplexy is a disorder in which an awake person collapses into a state of paralysis.
At the same time, the person may remain awake and have hypnogogic hallucinations
similar to dreaming.
KEY TERMS
REVIEW QUESTIONS
1. Why are circadian and circannual rhythms such prominent rhythms in
mammals?
2. Describe some of the details of the circadian clock that allow it to be easily
studied.
3. In what ways are NREM sleep and REM sleep organized differently in the brain?
4. Describe the various theories of sleep.
5. What are some of the most common sleep disorders, and what are their causes
and treatments?
FOR FURTHER THOUGHT
What ways can you suggest to combine the different theories of why we sleep into a
unified theory?
atonia, p. 458
basic rest–activity cycle
(BRAC), p. 464
beta rhythm, p. 455
biological clock, p. 442
biorhythm, p. 440
cataplexy, p. 477
circadian rhythm, p. 443
coma, p. 470
delta rhythm, p. 456
diurnal animal, p. 439
drug-dependency
insomnia, p. 474
entrainment, p. 445
free-running rhythm, p. 444
hypnogogic hallucination,
p. 477
insomnia, p. 473
locked-in syndrome,
p. 467
medial pontine reticular
formation (MPRF),
p. 472
melatonin, p. 453
microsleep, p. 467
narcolepsy, p. 473
non-REM (NREM) sleep,
p. 456
peribrachial area, p. 472
period, p. 442
place cell, p. 467
REM sleep, p. 456
reticular activating system
(RAS), p. 470
retinohypothalamic
pathway, p. 448
sleep apnea, p. 474
sleep paralysis, p. 477
slow-wave sleep, p. 456
suprachiasmatic nucleus
(SCN), p. 448
torpor, p. 439
Zeitgeber, p. 445
480 ! CHAPTER 12
neuroscience interact ive
Many resources are available for
expanding your learning on line:
www.worthpublishers.com/kolb/cha
pter12
Try some self-tests to reinforce your
mastery of the material. Look at some
of the updates on current research on
the brain. You’ll also be able to link to
other sites that will reinforce what
you’ve learned.
www.rls.org
Link to this site to learn more about
restless legs syndrome and current
research to help people suffering from
this disorder.
www.sleepfoundation.org
The headquarters of the National Sleep
Foundation can be a fascinating
starting point for investigation about
sleep disorders and normal sleep
patterns.
On your CD-ROM, you can review
some of the research methods useful to
understanding sleep in the threedimensional
research laboratory.
CH12.qxd 2/7/05 2:58 PM Page 480
ls
le
ll
RECOMMENDED READING
Dement,W. C. (1972). Some must watch while some must sleep. Stanford CA: Stanford
Alumni Association. This short book is written in an engaging style for the beginning
student of sleep. Nevertheless, instructors and even experts in sleep find it to be an
excellent introduction by one of the pioneers of sleep research.
Hobson, J. (1989). Sleep. New York: Scientific American Library. This book covers most of
the main ideas that have developed from research into sleep. It also covers areas of
psychology, ethology, neuroscience, and molecular biology, and it provides an overview
of the disorders of sleep. Hobson presents his own activation synthesis theory of
dreams.
Kleitman, N. (1965). Sleep and wakefulness. Chicago: University of Chicago Press. This book
is an exhaustive description of research into sleep and covers all the major findings
produced by the first decades of sleep research.
WHY DO WE SLEEP AND DREAM? ! 481
CH12.qxd 2/7/05 2:58 PM Page 481
Focus on New Research: Movement, Learning,
and Neuroplasticity
Connecting Learning and Memory
Studying Learning and Memory in the Laboratory
Two Categories of Memory
What Makes Explicit and Implicit Memory Different?
What Is Special about Personal Memories?
Dissociating Memory Circuits
Focus on Disorders: Patient Boswell’s Amnesia
Neural Systems Underlying Explicit
and Implicit Memories
Neural Circuit for Explicit Memories
Focus on Disorders: Alzheimer’s Disease
Focus on Disorders: Korsakoff’s Syndrome
Neural Circuit for Implicit Memories
Neural Circuit for Emotional Memories
Structural Basis of Brain Plasticity
Measuring Synaptic Change
Enriched Experience and Plasticity
Sensory or Motor Training and Plasticity
Plasticity, Hormones, Trophic Factors, and Drugs
Recovery from Brain Injury
Donna’s Experience with Brain Injury
Three-Legged Cat Solution
New-Circuit Solution
Lost-Neuron-Replacement Solution
482 !
C H A P T E R 13
How Do We Learn
and Remember?
Left: Oliver Meckes/Ottawa/Photo Researchers. Middle: Dimitri Messinis/
AP Photo. Right: ADEAR/RDF/Visuals Unlimited.
ls
le
ll
CH13.qxd 2/18/05 9:32 AM Page 482
Not all motor learning is good, however. Many musicians
develop focal hand dystonia, a condition in which
the person develops abnormal finger and hand positions,
cramps, and difficulty in coordinating hand and finger
movements. Dystonia can be so disabling that some musicians
must give up their occupation.
Typically, dystonia afflicts musicians who practice trying
to make perfect finger movements on their instruments.
Musicians at high risk include string players, who receive
vibratory stimulation at their fingertips. The constant practice
has been suggested to lead not only to improved musical
ability but also to distortion or disordering of the
motor maps in the cortex. Synchronous activation of the
digits by the vibration leads to this unwanted side effect.
Victor Candia and colleagues (2003) reasoned that the
dystonia was likely an example of disordered learning and
could be treated by retuning the motor map. The authors
used a neuroimaging technique known as magnetoencephalography
to measure changes in sensory-evoked magnetic
fields in the cortex.
At the beginning of the study, the musicians with dystonia
had a disordered motor map in which the fingers
overlapped one another, rather than being distinct. In training,
each subject used a hand splint tailored to his or her
individual hand anatomy. The splint allowed for the immobilization
of different fingers while the subjects made independent
movements of the others.
After 8 days of training for about 2 hours per day, the
subjects showed marked alleviation in the dystonic symptoms,
and the neuroimaging showed a normalization of the
cortical map with distinct fingers. Thus, training was able
to reverse the learned changes in the motor map and provide
a treatment for the dystonia. The musicians actually
“learned” a disorder, and they were able to “unlearn” it. In
view of the study by Ragert and colleagues, we can only
wonder whether the musicians’ unlearning was actually
faster because of the enhanced capacity to learn.
Movement, Learning, and Neuroplasticity
Focus on New Research
A s we travel through life, we are constantly changing
the organization of our brains by learning new
things. The neural basis for learning and memory resides at
the synapse, as detailed in Chapter 5. So, to learn anything
new, we must change the connections in the nervous system,
and, to do so, we keep adding new synapses.
What are the implications of all these changes? After
all, stated in Chapter 2, neural connections take up space.
Is there a limit to what we can learn? Does learning one
thing affect how we learn something else? If so, is the effect
good or bad? It can be both.
Many lines of research show that practicing motor
skills, such as playing a musical instrument, induces the organization
of the somatosensory and motor maps in the
cortex to change, as described in Chapter 10. The mental
maps generally become larger, at least for the finger and
hand representations. Presumably, musical skill improves,
but are other abilities enhanced, too?
Patrick Ragert and colleagues (2003) showed that professional
pianists have not only better motor skills but enhanced
somatosensory perception in their fingers as well.
When the researchers measured the ability to detect subtle
sensory stimulation of the fingertips, they found that the pianists
were more sensitive than controls.
They also found that the enhancement in tactile sensitivity
was related to the hours per day that the musicians
spent practicing. The investigators then asked whether the
enhanced perceptual ability precluded further improvement
in the musicians. Surprisingly, when both the musicians and
controls were given a 3-hour training session designed to
improve tactile sensitivity, the musicians showed more improvement
than did the controls, and, again, the extent of
improvement correlated with daily practice time.
This result implies that well-practiced musicians not
only learn to play music but also develop a greater capacity
for learning. Rather than using up all the available synapses,
they had developed a capacity to make even more.
ls
le
ll
CH13.qxd 2/18/05 9:32 AM Page 483
ls
le
ll
Most people tend to regard the brain as a relatively static structure that controls
behavior, but the brain changes throughout life, and these changes
allow us to modify our behavior, to adapt and learn, and to remember. In
Chapter 5, we introduced the term neuroplasticity to describe the nervous system’s potential
for changes that enhance its adaptability to environmental change and its ability
to compensate for injury. If we reflect on our own lives, we can easily compile a list
of experiences that must change the brain:
the profound changes during development discussed in Chapter 6;
the acquisition of culture;
preferences for certain foods or beverages or for art or other experiences; and
the ability to cope with neurodegeneration in the aging process and, for many, to
accommodate neurological injury or disease.
A characteristic common to all these examples is some form of learning. Understanding
how the brain supports learning is a fundamental question of neuroscience.
In Chapter 5, we analyzed different types of learning including habituation, sensitization,
and associative learning, at the level of the neuron.We noted in Experiment 5-4,
for example, that synapses change with events such as those observed in long-term potentiation
(LTP). Such changes can take place anywhere in the brain.
Exactly where in the brain might such changes take place when we learn specific
types of information? We can investigate the nature of the neural changes that support
learning by describing the changes in neurons exposed to specific sensory experiences.
Another strategy is to look at the neural changes that accompany various forms of
brain plasticity, such as recovery from brain injury or addiction to drugs. The overriding
goal of this chapter, however, is to generalize beyond learning to understand the nature
of behavior and the changing brain.
CONNECTING LEARNING AND MEMORY
Learning, as you know, is a change in an organism’s behavior as a result of experience.
Memory refers to the ability to recall or recognize previous experience. Memory thus
implies a mental representation of the previous experience. This hypothetical mental
representation is sometimes referred to as a memory trace, and neuroscientists presume
that a memory trace corresponds to some type of physical change in the brain.
At the macro level, however, what we know about the process of learning and the
formation of memories is inferred from changes in behavior, not observed directly. The
study of learning and memory therefore requires behavioral measures that evaluate
such behavioral changes.We begin here by looking at the ways that learning and memory
researchers study animals in the laboratory.We then look at what general types of
learning and memory can be identified from the results obtained in such studies.
Studying Learning and Memory in the Laboratory
A challenge for psychologists studying memory in laboratory animals (or people) is
to get the subjects to reveal what they can remember. Because laboratory animals do
not talk, investigators must devise ways for a subject to show its knowledge. Different
species can “talk” to us in different ways, and so the choice of test must be matched to
the capabilities of the species.
In the study of rats, mazes or swimming pools are typically used because rats
live in tunnels and near water. Studies of monkeys have taken advantage of the monkeys’
sharp vision and avid curiosity by requiring them to look under objects for
484 ! CHAPTER 13
Memory. The ability to recall or
recognize previous experience.
Pavlovian conditioning. Learning
procedure whereby a neutral stimulus
(such as a tone) comes to elicit a
response because of its repeated pairing
with some event (such as the delivery of
food); also called classical conditioning
or respondent conditioning.
Eye-blink conditioning. Commonly
used experimental technique in which
subjects learn to pair a formerly neutral
stimulus to a defensive blinking response.
Conditioned stimulus (CS). In
Pavlovian conditioning, an originally
neutral stimulus that, after association
with an unconditioned stimulus (UCS),
triggers a conditioned response.
Unconditioned stimulus (UCS). A
stimulus that unconditionally—naturally
and automatically—triggers a response.
Unconditioned response (UCR). In
classical conditioning, the unlearned,
naturally occurring response to the
unconditioned stimulus, such as
salivation when food is in the mouth.
Conditioned response (CR). In
Pavlovian conditioning, the learned
response to a formerly neutral
conditioned stimulus (CS).
Fear conditioning. Learned
association, a conditioned emotional
response, between a neutral stimulus and
a noxious event such as a shock.
CH13.qxd 2/18/05 9:32 AM Page 484
ls
le
ll
food or at television monitors. When birds
are the subjects, natural behaviors such as
singing are used. And, for human subjects,
there is a tendency to use paper-and-pencil
tests.
Consequently, psychologists have devised
hundreds of different tests in the past century
and, in doing so, have shown that there are
many types of learning and memory, each of
which appears to have its own neural circuitry.
Let us first consider some classic examples of
how animals can be trained to “talk” to investigators.
A century ago, two very different traditions
emerged for studying learning and
memory, one based on the work of Edward
Thorndike in the United States and the other
on experiments conducted by Ivan Pavlov in
Russia.
PAVLOVIAN CONDITIONING
In the early years of the twentieth century, Russian physiologist Ivan Pavlov discovered
that, when a food reward accompanied some stimulus, such as a tone, dogs learned to
associate the stimulus with the food. Then, whenever they heard the tone, they would
salivate even though no food was present. This type of learning has many names, including
Pavlovian conditioning, respondent conditioning, and classical conditioning,
and its characteristics have been documented by many studies.
A key feature of Pavlovian conditioning is that animals learn the association between
two stimuli (such as the presentation of the food and the tone) and tell us that
they have learned it by giving the same response (such as salivation) to both stimuli.
Pet owners are familiar with this type of learning: to a cat or dog, the sound of a can
opener is a clear stimulus for food. Experimentally, two forms of Pavlovian conditioning
are commonly used today: eye-blink conditioning and fear conditioning. These
forms have proved especially useful because they are associated with neural circuits in
discrete brain regions.
Eye-blink conditioning has been used to study Pavlovian learning in rabbits
and people (Figure 13-1). In these studies, a tone (or some other stimulus) is associated
with a painless puff of air to the eye of the subject. The tone, which is the conditioned
stimulus (CS), comes to elicit a blink that is initially produced by the air puff, which
is the unconditioned stimulus (UCS), because blinking is the normal reaction—the
unconditioned response (UCR)—to a puff of air. Thus, the subject tells us that it has
learned that the signal stimulus predicts the puff by blinking in response to the signal
alone—a conditioned response (CR).
This form of learning is mediated by circuits in the cerebellum (see Figure 10-14).
The cerebellum does not have special circuits just for eye-blink conditioning, which
is an artificial situation. Rather, the cerebellum has circuits designed to pair various
motor responses with environmental events. Eye-blink conditioning experiments simply
take advantage of this biological predisposition.
In fear conditioning, a noxious stimulus is used to elicit fear, an emotional response.
A rat or other animal is placed in a box that has a grid floor through which a
mild but noxious electric current can be passed. As shown in Experiment 13-1, a tone
(the CS) is presented just before a brief, unexpected, mild electric shock. (This shock
is roughly equivalent to the static-electrical shock that we get when we rub our feet on
a carpet and then touch a metal object or another person.)
HOW DO WE LEARN AND REMEMBER? ! 485
Headgear is
arranged for eyeblink
conditioning.
1
Puff of air to
eye causes eye
to blink.
2
After pairing air puff
with tone, tone alone
comes to elicit a blink.
3
Electrodes
Air jet
tube
Audio
speaker
Figure 13-1
Eye-Blink Conditioning A subject
wears headgear containing an apparatus
that delivers a puff of air to the eye,
which causes the subject to blink. When
the air puff is paired with a tone, the
subject learns the association and
subsequently blinks to the tone alone.
Circuits in the cerebellum mediate this
form of learning.
Later
Rat is given mild electrical shock
in combination with tone.
The rat has learned an association
between the tone and the shock, which
produces a fear response. Circuits that
include the amygdala take part in this
learning process.
Conclusion
Question: Does an animal learn the association
between emotional experience and environmental
stimuli?
EXPERIMENT 13-1
Later
Rat freezes in fear when tone
is given alone.
If a light is presented alone
later, rat ignores it.
Tone
plus shock
Light only—
no tone
Tone only
Procedure and results
CH13.qxd 2/18/05 9:32 AM Page 485
ls
le
ll
When the tone is later presented without the shock, the animal will act afraid. It may
become motionless and may urinate in anticipation of the shock. Presentation of a novel
stimulus, such as a light, in the same environment has little effect on the animal. Thus,
the animal tells us that it has learned the association between the tone and the shock.
Because the CR is emotional, circuits of the amygdala, rather than the cerebellum,
mediate fear conditioning. Although both eye-blink and fear conditioning are Pavlovian,
different parts of the brain mediate the learning.
INSTRUMENTAL CONDITIONING
The second tradition of studying learning and memory was begun in the United States
by Edward Thorndike (1898). Thorndike was interested in how animals solve problems.
In one series of experiments, Thorndike placed cats in a box with a plate of fish
outside it (Figure 13-2). The only way for a hungry cat to get to the fish was to figure
out how to get out of the box.
The solution was to press on a lever, which activated a system of pulleys that
opened the box’s door. The cat gradually learned that its actions had consequences: on
the initial trial, the cat touched the releasing mechanism only by chance as it restlessly
paced inside the box. The cat apparently learned that something that it had done
opened the door, and it tended to repeat the behaviors that had occurred just before
the door opening. After a few trials, the cat took just seconds to get the door open so
that it could devour the fish.
Later studies by B. F. Skinner (e.g., 1938) used a similar strategy of reinforcement
to train rats to press bars or pigeons to peck keys to obtain food (Chapter 11). Many
animals will learn to bar press or key peck if they are simply placed into the apparatus
and allowed to discover the response necessary to obtain the reward, just as Thorndike’s
cats learned to escape his puzzle boxes. This type of learning is instrumental conditioning,
or operant conditioning. The subject demonstrates that it has learned the association
between its actions and the consequences by the increasing speed at which it
can perform the task.
The variety of such instrumental associations is staggering, as we are constantly
learning the association between our behavior and its consequences. It should be no
surprise, therefore, that instrumental learning is not localized to any particular circuit
in the brain. The circuits needed vary with the actual requirements of the task, as is
demonstrated by the following examples.
Richard Morris devised a task in 1980 that has become popular in research in
learning and memory.He placed rats in a large swimming pool in which an escape platform
was invisible to the rats because it was just under the water’s surface. (Figure 13-
486 ! CHAPTER 13
Lever
Pulley
system
Although learning is not
immediate, the hungry
cat eventually learns
that pressing on the
lever will result in
getting out of the box
and being able to reach
the food.
The cat is placed in the box
with the food reward outside.
Food
reward
Figure 13-2
Thorndike’s Puzzle Box
Instrumental conditioning. Learning
procedure in which the consequences
(such as obtaining a reward) of a
particular behavior (such as pressing a
bar) increase or decrease the probability
of the behavior occurring again; also
called operant conditioning.
Visuospatial learning. Using visual
information to identify an object’s
location in space.
CH13.qxd 2/18/05 9:32 AM Page 486
ls
le
ll
HOW DO WE LEARN AND REMEMBER? ! 487
(A)
(C) Matching-to-place learning
T1
T2
(B) Place learning
T1
T5
T4
T3
T2
Pool
Hidden
platform
T = Trial
Submerged
platform
Room
cues
(D) Landmark learning
T1
T3
T2
Cue
(on wall
of pool)
Figure 13-3
Instrumental Conditioning (A) General arrangement of the swimming pool used in three
different visuospatial learning tasks for rats. The red lines in parts B, C, and D mark the rat’s
swimming path on each trial (T). (B) In a place-learning task (Morris, 1981), a rat is put into the
pool at various starting locations and must learn the location of a hidden platform, which can
be done only by considering the configuration of visual cues in the room—windows, wall
decorations, potted plants, and the like. (C) In a matching-to-place task (Whishaw, 1989), the
rat is again put in the pool at random locations, but the hidden platform is in a new location
each test day. The animal must learn that the location where it finds the platform on the first
trial of each day is the location of the platform for all that day’s trials. (D) In a landmarklearning
task (Kolb & Walkey, 1987), the rat is required to ignore the room cues and to learn
that only the cue on the wall of the pool signals the location of the platform. The platform
and cue are moved on each trial so that the animal is penalized for trying to use room cues to
solve the problem.
3A shows the setup.) The task for the rat was to discover that there was an escape from
the water and then to figure out where the platform was.
In one version of the test, illustrated in Figure 13-3B, the only available cues were
distal ones—that is, external to the pool. Because no single cue would identify the location
of the platform, the rats had to learn the relation between several cues in the
room and the platform’s location. To do so obviously requires visuospatial learning,
the use of visual information to identify an object’s location in space.
Rats normally learn the Morris task in just a few trials such that, when placed anywhere
in the pool, they can swim directly to the hidden platform.We can infer that a
rat that is able to swim but is unable to learn this task has some disturbance in the neural
circuits underlying visuospatial learning.
In a variation of the task, after rats have been trained to find the platform, its location
is changed (Figure 13-3C). The platform is moved to a new position every day; the
rats must find the platform on the first trial in the pool and then go to that location for
the remainder of that day’s trials. In this case, the rats’ task is to learn not only the platform’s
location with respect to the visual world but also its new location each day.
For a simulation on learning, go to the
Web site at www.worthpublishers.
com/kolb/chapter13
CH13.qxd 2/18/05 9:32 AM Page 487
ls
le
ll
Rats quickly learn this puzzle and form what is known as a learning set, an understanding
of how a problem can be solved through the use of a rule that can be applied
in many different situations. In the current example, the rule is that the successful solution
(finding the platform) requires a shift in strategy when the old strategy fails.Welltrained
animals need only a single trial to learn the platform’s location each day, and
they will swim flawlessly to that location on subsequent trials. This type of task, in which
the rat must keep track of a specific piece of information on a given day, places demands
on the brain that are clearly different from the simpler Morris version, in which the
learning is gradual, much like that in Thorndike’s cats.
In yet another variation, a cue can be placed on the wall of the pool (Figure
13-3D). The rat’s task is to learn that the cue, and only the cue, indicates the approximate
location of the platform. In this case, the platform moves on every trial but always
maintains the same relation to the cue, which also moves. The brain therefore is
learning that all the distal cues are irrelevant and only the local cue is relevant. This
task is very different, and, once again, different neural circuitry is required to solve it.
Two Categories of Memory
Humans present a different challenge to the study of memory because so much of our
learning is verbal. Psychologists have been studying human memory since the mid-
1800s, and cognitive psychologists have developed sophisticated measures of learning
and memory for neuropsychological investigations. Two such measures will help us
distinguish between two categories of memory in humans.
In one kind of task, a group of subjects are given a list of words to read, such as spring,
winter, car, and boat. Another group of subjects read a list consisting of the words trip,
tumble, run, and sun. All the subjects are then asked to define a series of words, one of
which is fall. The word fall has multiple meanings, including the season and a tumble.
People who have just read the word list containing names of seasons are likely to give the
“season” meaning, whereas those who have read
the second list, containing action words, will give
the “tumble”meaning. Some form of unconscious
(and unintentional) learning takes place as the
subjects read the word lists.
This word-list task is a measure of implicit
memory. People with amnesia, a partial or total
loss ofmemory,perform normally on such tests of
implicit memory. The amnesic person has no recollection
of having read the word list yet acts as
though some neural circuit has been influenced by
it. Thus, there is a dissociation between the memory
of the unconscious (or implicit) learning and
the conscious recollection of training,which is referred
to as explicit memory.
This implicit–explicit distinction is not restricted
to verbal learning but is true of visual
learning and motor learning tasks as well. For example,
subjects can be shown the top panel of the
Gollin figure test in Figure 13-4 and asked what it
shows. They are unlikely to be able to identify an
image, and so they are presented with a succession
of more nearly complete sketches until they can
identify the picture.
488 ! CHAPTER 13
Learning set. An understanding of how
a problem can be solved with a rule that
can be applied in many different
situations.
Implicit memory. Memory in which
subjects can demonstrate knowledge but
cannot explicitly retrieve the information.
Amnesia. Partial or total loss of memory.
Explicit memory. Memory in which a
subject can retrieve an item and indicate
that he or she knows that the retreived
item is the correct item (i.e., conscious
memory).
Figure 13-4
Gollin Figure Test
Subjects are shown a series
of drawings in sequence,
from least to most clear,
and asked to identify the
image. The object cannot
be identified in the first
sketch, and most people
must see several of the
panels before they can
identify it. On a retention
test some time later,
however, subjects identify
the image sooner than they
did on the first test,
indicating some form of
memory for the image.
Amnesic subjects also show
improvement on this test,
even though they do not
recall taking the test
before.
CH13.qxd 2/18/05 9:32 AM Page 488
ls
le
ll
When control subjects and amnesics are later shown the same sketch, both groups
are able to identify the figure sooner than they could the first time. Even though the
amnesic subjects may not recall having seen the sketches before, they behave as though
they had.
A second kind of measure reveals implicit learning of motor skills. For instance, a
person can be taught a motor skill, such as the pursuit-rotor task shown in Figure 13-5.
A small metal disc moves in a circular pattern on a turntable that also is moving. The
task is to hold a stylus on the small disc as it spins. This task is not as easy as it looks,
especially when the turntable is moving quickly.
Nonetheless, with an hour’s practice,most people become reasonably proficient. If
they are presented with the same task a week later, both normal subjects and amnesics
will take less time to perform it. Here, too, the amnesics will fail to recall having ever
performed the task before.We can see, therefore, that the distinction between tests of
implicit and explicit memory is consistent and must provide
some key to how the brain stores information.
The implicit–explicit distinction is just one way in
which psychologists have categorized different memory
processes. Many researchers prefer to distinguish between
declarative memory, which refers to the specific contents
of specific experiences that can be verbally recalled, such
as times, places, or circumstances, and procedural memory,
which is the ability to to do something.Although some theorists
may make subtle distinctions between the implicit–
explicit and declarative–procedural dichotomies, as applied
to humans there is really little practical difference and we
will use the explicit–implicit dichotomy.
Table 13-1 lists other commonly used dichotomies,
with the general distinction being that one memory category
requires the recollection of specific information
whereas the other refers to knowledge of which we are not
consciously aware.We can include Pavlov’s classical conditioning,
Thorndike’s instrumental learning, and Skinner’s
operant learning in this analysis, too, because all of them
are types of implicit learning.
Nonspeaking animals can display explicit memory.
One of us owned a cat that loved to play with a little ball.
One day the ball was temporarily put on a high shelf to keep
HOW DO WE LEARN AND REMEMBER? ! 489
Declarative memory. Ability to recount
what one knows, to detail the time, place,
and circumstances of events; often lost in
amnesia.
Procedural memory. Ability to recall a
movement sequence or how to perform
some act or behavior.
Rotating
target Stylus
Rotating
disk
Figure 13-5
Pursuit-Rotor Task The subject must keep the stylus in contact with
the metal disc that is moving in a circular pattern on a turntable,
which also is moving in a circular pattern. Although the task is
difficult, most people show significant improvement after a brief
period of training. When given a second test at some later time, both
normal subjects and amnesics show retention of the task. The
amnesics typically do not recall learning the task before.
Terms Describing Two Categories of Memory
Term for conscious memory Term for unconscious memory
Explicit Implicit
Declarative Procedural
Fact Skill
Memory Habit
Knowing that Knowing how
Locale Taxon
Cognitive mediation Semantic
Conscious recollection Skills
Elaboration Integration
Memory with record Memory without record
Autobiographical Perceptual
Representational Dispositional
Episodic Semantic
Working Reference
Note: These paired terms have been used by various theorists to differentiate conscious
from unconscious forms of memory. This list is intended to help you relate other
discussions of memory that you may encounter to the one in this book, which favors
the explicit–implicit distinction.
Table 13-1
CH13.qxd 2/18/05 9:32 AM Page 489
ls
le
ll
it away from an inquisitive 1-year-old boy. For weeks afterward, the cat would sit and
stare at the location where the ball had been placed—an example of explicit memory.
Animals also display explicit memory when they learn psychological tasks. Recall
that, in one variant of the Morris task, rats were given a new platform location on each
day of training. The task therefore was to go to the platform’s last location. This piece
of information is explicit and can be demonstrably forgotten.
Suppose that a well-trained rat is given one trial with the platform at a new location
and then not given a second trial for an hour, a day, 3 days, or a week. The rat has
no difficulty with a delay of an hour or even a day. Some rats are flawless at 3 days, but
most have forgotten the location by the time a week has elapsed. Instead, they swim
around looking for the platform. This behavior illustrates their implicit memory of the
learning set, or the “rules of the game”—namely, that there is a platform and that it can
be found with a certain type of search strategy.
What Makes Explicit and Implicit
Memory Different?
One reason that explicit and implicit memories differ is that each type of memory is
housed in a different set of neural structures. Another reason that they differ is that explicit
and implicit information are processed differently.
ENCODING MEMORIES
Implicit information is encoded in very much the same way as it is perceived and can
be described as data-driven, or “bottom up,” processing. The idea is that information
enters the brain through the sensory receptors and is then processed in a series of
subcortical and cortical regions. For example, recall from Chapter 8 that visual information
about an object goes from the visual receptors (the “bottom”) to the lateral
geniculate nucleus, the occipital cortex, and then through the ventral stream to the
temporal lobe, where the object is recognized.
Explicit memory, in contrast, depends on conceptually driven, or “top down,” processing,
in which the subject reorganizes the data. For example, if you were searching
for a particular object such as your set of keys, you would ignore other objects. This is
a top-down process because circuits in the temporal lobe (the “top”) form an image
that influences how incoming visual information (the “bottom”) is processed, which
in turn greatly influences the recall of information later.
Because a person has a relatively passive role in encoding implicit memory, he or
she will have difficulty recalling the memory spontaneously but will recall the memory
more easily when primed by the original stimulus or some feature of it. Because a person
plays an active role in processing information explicitly, the internal cues that were
used in processing can also be used to initiate spontaneous recall.
Findings from studies of eyewitness testimony demonstrate the active nature of recalling
an explicit memory and its potential fallability (e.g., Loftus, 1997). In a typical
experiment, people are shown a video clip of an accident in which a car collides with
another car stopped at an intersection. One group of subjects is asked to estimate how
fast the car was going when it “smashed” into the other car.A second group is asked how
fast the car was going when it “bumped” into the other car.
Later questioning indicates that the memory of how fast the first car was moving
is biased by the instruction: subjects looking for “smashing” cars estimate faster speeds
than those estimated by subjects looking for “bumping” cars. In other words, the instruction
causes the information to be processed differently. In both cases, the subjects
were certain that their memories were accurate.
490 ! CHAPTER 13
Priming. Using a stimulus to sensitize
the nervous system to a later presentation
of the same or a similar stimulus.
To test your own memory, go to the
Web site at www.worthpublishers.
com/kolb/chapter13
CH13.qxd 2/18/05 9:32 AM Page 490
ls
le
ll
Other experiments show that implicit memory also is fallible. For example, subjects
are read the following list of words: sweet, chocolate, shoe, table, candy, horse, car,
cake, coffee, wall, book, cookie, hat. After a delay of a few minutes, the subjects hear another
list of words that includes some of the words from the first list and some that are
new. Subjects are asked to identify which words were present on the first list and to indicate
how certain they are of the identification.
One of the words on the second list is sugar. Most subjects indicate not just that
sugar was on the first list but that they are certain that it was.Although other sweet things
were, sugar was not. This type of demonstration is intriguing, because it shows the ease
with which we can form “false memories” and defend their veracity with certainty.
PROCESSING MEMORIES
Although memories can be distinguished generally as implicit or explicit, the brain
does not process all implicit or all explicit memories in the same way. Memories can
be divided according to categories that are different from those listed in Table 13-1. For
example, we can make a distinction between memories for different types of sensory
information.
We have seen that visual and auditory information is processed by different neural
areas, and so it is reasonable to assume that auditory memories are stored in different
brain regions from those in which visual memories are stored.We can also make a distinction
between information stored in so-called short-term memory and information
held for a longer time in long-term memory. In short-term memory, information—such
as the telephone number of a restaurant that we have just looked up in the Yellow
Pages—is held in memory only briefly, for a few minutes at most, and then is discarded.
In long-term memory, information—such as a close friend’s name—is held in memory
indefinitely, perhaps for a lifetime.
The frontal lobes play an important role in short-term memory, whereas the temporal
lobe plays a central role in long-term storage of verbal information. The crucial
point is that no single place in the nervous system can be identified as the location for
memory or learning. Virtually the entire nervous system can be changed with experience,
but different parts of an experience change different parts of the nervous system.
One challenge for the experimenter is to devise ways of manipulating experience to
demonstrate change in different parts of the brain.
STORING MEMORIES
Acceptance of the idea that every part of the brain can learn influences our view of the
nature of the neural circuits that mediate memory. For one thing, we could expect that
areas that process information also house the memory of that information. Areas that
process visual information, for example, likely house visual memory. Because the temporal
lobe has specialized regions for processing color, shape, and other visual information
regarding an object’s characteristics, we can predict that the memory for
various visual attributes of objects will be stored separately.
This prediction has been confirmed by a series of PET studies by Alex Martin and
colleagues (1995) at the U.S. National Institute of Mental Health. In one of these studies,
subjects were shown black-and-white line drawings of objects and asked to generate
words denoting either colors or actions of the objects. The idea was that the
processing of color and motion is carried out in different locations in the temporal
lobe, and thus the activity associated with the memories of color and motion also
might be dissociated.
In fact, just such a dissociation was demonstrated. Figure 13-6 shows that recall of
colors activates a region in the ventral temporal lobe, just anterior to the area controlling
color perception, whereas recall of action words activates a region in the middle
HOW DO WE LEARN AND REMEMBER? ! 491
KEY
Color words
Action words
Overlap
Middle temporal gyrus
Inferior temporal gyrus
Superior temporal gyrus
Frontal
activation
Parietal
activation
Temporal
activation
Figure 13-6
Memory Distribution Lateral view of
the left hemisphere shows regions of
increased blood flow when subjects
generate color words (red) and action
words (blue) to describe objects shown to
them in static, black-and-white drawings.
Purple indicates areas of overlap. The
red region extends under the lateral part
of the temporal lobe. These data suggest
that object memory is organized as a
distributed system in which the attributes
of an object are stored close to the
regions of the cortex that mediate the
perception of those attributes. Activation
in the parietal lobe is likely related
to the movements associated with
actions, and in the frontal lobe to the
spontaneous generation of behavior.
Adapted from “Discrete Cortical Regions
Associated with Knowledge of Color and
Knowledge of Action,” by A. Martin, J. V. Haxby,
F. M. Lalonde, C. L. Wiggs, and L. G. Ungerleider,
1995, Science, 270, p. 104.
CH13.qxd 2/18/05 9:32 AM Page 491
ls
le
ll
temporal gyrus, just anterior to the area controlling the perception of motion. This distribution
of neural activation shows not only that object memory is at least partly located
in the temporal lobe but also that it is found in regions associated with the
original perception of the objects.
What Is Special about Personal Memories?
One aspect of memory that is unique to each of us is our personal, or autobiographical,
memory. This episodic memory includes not only a record of events (episodes)
that took place but also our presence there and our role in the events. Our personal experiences
form the basis of who we are and the rules by which we live. That is, we have
memories not only for events but also for their context at a particular time in a particular
place. We thus have a concept of time and a sense of our personal role in a
changing world.
Imagine what would happen if we lost our personal memory? We would still recall
events that took place, but we would be unable to see our role in them. People with
frontal-lobe injuries sometimes exhibit such symptoms, as illustrated in a case described
by Endel Tulving (2002).
K. C. had a serious traumatic brain injury in a motorcycle accident that produced
multiple cortical and subcortical lesions.What is remarkable about K. C., however, is
that his cognitive abilities are intact and indistinguishable from most normal healthy
adults. He can still play chess and the organ, and his short-term memory is intact. He
knows who he is, when his birthday is, the names of schools that he attended, the location
of the family cottage, and so on.
What K. C. cannot do is recall any personally experienced events. This episodic
amnesia covers his entire life, from birth until the present. Thus, whereas he knows
facts about himself, he has no memory for events that included him personally. For example,
he is unable to describe an event that took place in school that specifically included
him, while at the same time recalling going to school and the knowledge that
he gained there.
Findings from neuroimaging studies of people with episodic amnesia suggest that
they consistently have frontal-lobe injuries (Lepage et al., 2000), but exactly why these
lesions produce episodic amnesia is still unclear. Nonetheless, Tulving made the interesting
proposal that episodic memory is a marvel of nature: it transforms the brain into
a kind of time machine that allows us to dwell on the past and make plans for the future.
He goes further to suggest that this ability may be unique to humans and is presumably
due to some novel evolutionary development of the frontal lobe.
Not all people with episodic amnesia have brain injury, however.Many case reports
describe patients with massive memory disturbances resulting
from some sort of “psychiatric” or “psychogenic”disorder.
Such cases have been fodder for numerous movie plots.
Hans Markowitsch (2003) noted that the amnesia reported
in some of these cases is remarkably similar to episodic
amnesia seen in neurological patients.Neuroimaging of
patients with psychogenic amnesia shows a massive reduction
in brain activity in frontal regions that is remarkably similar
to that seen in neurological patients with episodic amnesia
(Figure 13-7). Therefore patients with psychogenic amnesias
can be assumed to have a dysfunction of frontal-brain
activity that acts to block the retrieval of autobiographical
memory.
492 ! CHAPTER 13
Episodic memory. Autobiographical
memory for events pegged to specific
place and time contexts.
Figure 13-7
Lost Episodes Horizontal images
through the brains of two patients
with selective retrograde amnesia for
autobiographical information. The
section on the left is from an amnesic
patient who had a brain infection
(herpes simplex encephalitis). The right
frontal and temporal lobes are dark,
owing to a metabolic reduction in the
right temporofrontal region (arrow). The
section on the right shows the brain of
a patient with psychogenic amnesia.
Again, a significant metabolic reduction
is visible in the right temporofrontal
area (arrow). From “Functional Neuroimaging
Correlates of Functional Amnesia,” by H. J.
Markowitsch, 1999, Memory 7, Plate 2. Reprinted
by permission of Psychology Press Ltd., Hove.
CH13.qxd 2/18/05 9:32 AM Page 492
ls
le
ll
DISSOCIATING MEMORY CIRCUITS
Beginning in the 1920s and continuing until the early 1950s, American psychologist
Karl Lashley looked for the neural circuits underlying memory for the solutions to
mazes learned by laboratory rats and monkeys. Lashley’s working hypothesis was that
memories must be represented in the perceptual and motor circuitry used to learn solutions
to problems.He believed that, if he removed bits of this circuitry or made knife
cuts that disconnected it, amnesia should result.
In fact, however, neither procedure produced amnesia.
Lashley found that the severity of the memory disturbance
was related to the size of the injury rather than to its
location. In 1951, after 30 years of searching, Lashley concluded
that he had failed to find the location of the memory
trace, although he did believe that he knew where it was
not located (Lashley, 1960).
Ironically, just two years later William Scoville made a
serendipitous discovery that had not been predicted by
Lashley’s studies. On August 23, 1953, Scoville performed a
bilateral medial-temporal-lobe resection on a young man
who became known as Case H.M.H.M. had severe epilepsy
that was not controlled by medication. Like Wilder Penfield
(see Chapter 9), Scoville was a neurosurgeon who was
attempting to rid people of seizures by removing the abnormal
brain tissue that was causing them.
H. M.’s seizures were originating in the medial temporal
region, which includes the amygdala, hippocampal
formation, and associated cortical structures, and so Scoville
removed them bilaterally, leaving the more lateral
temporal lobe tissue intact. As shown in Figure 13-8, the
removal included the anterior part of the hippocampus,
In Review .
Among the multiple forms of learning, a process that results in a relatively permanent
change in behavior as a result of experience, a primary distinction can be made between
Pavlovian conditioning, in which some environmental stimulus (such as a tone) is paired
with a reward, and operant conditioning, in which a response (such as pushing a button)
is paired with a reward. The demands on the nervous system are different in the two types
of learning, and so we can expect that the regions of the brain related to each learning
form will be different.
Memory is the ability to recall or recognize previous experience; this definition implies
the existence of a memory trace, or a mental representation of a previous experience.
There are many forms of memory, each related to mental representations in different parts
of the brain. One useful distinction is between implicit memory, in which information is
unconsciously learned, and explicit memory, which is conscious memory for specific information.
The mental representations of implicit and explicit memory are held in different
regions of the brain. Another form of memory, episodic memory, is autobiographical
and unique to each person.
HOW DO WE LEARN AND REMEMBER? ! 493
(A)
(B)
(C)
Area of
lesion
Entorhinal cortex Amygdala
Entorhinal
cortex
Collateral
sulcus
Hippocampus
Hippocampus
Figure 13-8
Extent of H. M.’s Surgery H. M.’s brain viewed ventrally, with the righthemisphere
lesion highlighted. The left side of the brain has been left intact
to show the relative location of the medial temporal structures. Because the
lesion runs along the wall of the medial temporal lobe, it can be seen in
several cross sections of the brain. Parts A, B, and C, based on MRI scans,
depict such sections of H. M.’s brain. Adapted from “H. M.’s Medial Temporal Lobe
Lesion: Findings from Magnetic Resonance Imaging,” by S. Corkin, D. G. Amaral, R. G.
Gonzalez , K. A. Johnson, and B. T. Hyman, 1997, Journal of Neuroscience, 17, p. 3966.
CH13.qxd 2/18/05 9:32 AM Page 493
ls
le
ll
the amygdala, and the adjacent cortex. The behavioral symptoms that Scoville noted
after the second surgery were completely unexpected, and so he invited Brenda Milner,
one of Penfield’s associates, to study H. M. Milner and her colleagues have studied H.
M. for more than 50 years, making him the most studied case in neuroscience (e.g.,
Corkin, 2002).
H. M.’s most remarkable symptom is severe amnesia: he is unable to recall anything
that has happened since his surgery in 1953.H.M. still has an above-average I.Q.
score (118 on the Wechsler Adult Intelligence Scale; 100 is average), and he performs
normally on perceptual tests. Furthermore, his recall of events from his childhood and
school days is intact. Socially, H. M. is well mannered, and he can engage in sophisticated
conversations. However, he cannot recall events that have just happened. H. M.
has no explicit memory.
In one study by Suzanne Corkin,H.M. was given a tray of hospital food, which he
ate. A few minutes later, he was given another tray. He did not recall having eaten the
first meal and proceeded to eat another. A third tray was brought, and this time he ate
only the dessert, complaining that he did not seem to be very hungry.
To understand the implications and severity of H. M.’s condition, one need only
consider a few events in his postsurgical life. His father died, but H. M. continued to
ask where his father was, only to experience anew the grief of hearing that his father
had passed away. (Eventually H. M. stopped asking about his father, suggesting that
some type of learning had taken place.)
Similarly, when in the hospital, he typically asks, with many apologies, if the nurses
can tell him where he is and how he came to be there. He remarked on one occasion,
“Every day is alone in itself, whatever enjoyment I’ve had, and whatever sorrow I’ve
had.”His experience is that of a person who perceives his surroundings but cannot comprehend
the situation that he is in because he does not remember what has gone before.
Formal tests of H.M.’s memory show what one would expect: he cannot recall specific
information just presented. In contrast, his implicit-memory performance is
nearly intact. He performs normally on tests such as the incomplete-figure or pursuitrotor
tasks illustrated in Figures 13-4 and 13-5.Whatever systems are required for implicit
memory must therefore be intact, but those systems crucial to explicit memory
are misssing or dysfunctional. Another case, similar to that of H. M., is discussed in
“Patient Boswell’s Amnesia.” Curiously, H. M. can recognize faces, including his own
face, and he recognizes that he has aged. Face recognition depends on the parahippocampal
gyrus, which is partly intact on H. M.’s right side.
There are probably several reasons why Lashley did not find a syndrome like that
shown by H. M. Most important, Lashley did not use tests of explicit memory, and so
his animal subjects would not have shown H. M.’s deficits. Rather, Lashley’s tests were
mostly measures of implicit memory, with which H.M. has no problems. The following
case illustrates that Lashley probably should have been looking in the basal ganglia
for deficits revealed by his tests of implicit memory.
J. K. was born on June 28, 1914. He was above average in intelligence and worked
as a petroleum engineer for 45 years. In his mid-70s, he began to show symptoms of
Parkinson’s disease, and, at about age 78, he started to have memory difficulties. (Recall
that, in Parkinson’s disease, the projections from the dopaminergic cells of the
brainstem to the basal ganglia die.)
Curiously, J. K.’s memory disturbance was related to tasks that he had done all his
life. On one occasion, he stood at the door of his bedroom frustrated by his inability
to recall how to turn on the lights. He remarked, “I must be crazy. I’ve done this all my
life and now I can’t remember how to do it!” On another occasion, he was seen trying
to turn the radio off with the television’s remote control. This time he explained, “I
don’t recall how to turn off the radio so I thought I would try this thing!”
494 ! CHAPTER 13
CH13.qxd 2/18/05 9:32 AM Page 494
ls
le
ll
J. K. clearly had a deficit in implicit memory. In contrast, he was aware of daily
events and could recall explicit events as well as most men his age.He could still speak
intelligently on issues of the day that he had just read about. Once when we visited
him, one of us entered the room first and he immediately asked where the other
was, even though it had been 2 weeks since we told him that we would be coming to
visit.
This intact long-term memory is very different from the situation of H. M., who
would not have remembered that anybody was coming, even 5 min after being told.
Because Parkinson’s disease primarily affects the basal ganglia, J. K.’s deficit in implicit
memory was likely related to his basal ganglia dysfunction.
HOW DO WE LEARN AND REMEMBER? ! 495
Patient Boswell’s Amnesia
Focus on Disorders
Boswell is a man who, at the age of 48, developed a brain
infection known as herpes simplex encephalitis. Before his
illness, Boswell had had 13 years of schooling and had
worked for nearly 30 years in the newspaper advertising
business. By all accounts, he was successful in his profession
and was a normal, well-adjusted person.
Boswell recovered from the acute symptoms of the disease,
which included seizures and a 3-day coma. His postdisease
intelligence was low average, probably owing to
the neurological damage caused by the disease. Nonetheless,
his speech and language remained normal in every respect,
and he suffered no defects of sensory perception or of
movement.
But Boswell was left with a severe amnesic syndrome. If
he hears a short paragraph and is asked to describe its main
points, he routinely gets scores of zero. He can only guess
the day’s date and is unable even to guess what year it is.
When asked what city he is in, he simply guesses. He does
know his place of birth, and he can correctly recall his birth
date about half the time. In sum, Boswell has a severe amnesia
for events both before and since his encephalitis. He
does show implicit memory, however, on tests such as the
pursuit-rotor task (see Figure 13-5).
Boswell’s amnesia has been extensively investigated by
Antonio Damasio and his colleagues (1989), and his brain
pathology is now well documented. The critical damage is a
bilateral destruction of the medial temporal regions and a
loss of the basal forebrain and the posterior part of the orbital
frontal cortex. In addition, Boswell has lost the insular
cortex, which is found in the lateral fissure and not visible in
the adjoining illustration.
In contrast, his sensory and motor cortices are intact, as
are his basal ganglia. Boswell’s injury is thus more extensive
than H. M.’s. Like H. M., he has a loss of new memories, but,
unlike H. M., he also has a severe loss of access to old information,
probably because of his insular and prefrontal injuries.
Nonetheless, again like H. M., Boswell has an intact
procedural memory, a fact that illustrates the dissociation between
neural circuits underlying explicit and implicit forms
of memory.
After a herpes simplex encephalitis infection, patient Boswell has
severe amnesia and has difficulty remembering events before and
after his illness. This model highlights the areas of damage in the
medial temporal region, the basal forebrain, and the posterior
orbital frontal cortex. Compare Figure 13-7.
Lateral view
Left hemisphere Right hemisphere
Medial view
Damaged
area
Damaged
area
CH13.qxd 2/18/05 9:32 AM Page 495
ls
le
ll
NEURAL SYSTEMS UNDERLYING EXPLICIT AND
IMPLICIT MEMORIES
Findings from laboratory studies, largely on rats and monkeys, have shown that the
symptoms of patients such as H. M. and J. K. can be reproduced in animals by injuring
the medial temporal region and basal ganglia, respectively. Other structures, most
notably in the frontal and temporal lobes, also have been found to play roles in certain
types of explicit memory.We now consider the systems for explicit and implicit memory
separately.
Neural Circuit for Explicit Memories
The dramatic amnesic syndrome discovered in H. M. in the 1950s led investigators to
focus on the hippocampus, which at the time was regarded as a large brain structure
in search of a function. However, because H. M. has damage to other structures, too,
the initial view that the hippocampus is the location of explicit-memory processing
turned out to be incorrect. It took several decades of anatomical and behavioral studies
to sort out the complexities, but, by the mid-1990s, a consensus began to emerge on
a theory for explicit memory. Note that, if you consult books or reviews published before
then, the explanation may be quite different (see Gazzaniga, 2000).
The prime candidates for a role in explicit memory include the medial temporal
region, the frontal cortex, and structures closely related to them. Before considering the
model, we must first revisit the anatomy of the medial temporal region. As we do so, it
is important to keep in mind the studies by Martin and colleagues, discussed earlier
(look again at Figure 13-6). Findings from those studies showed that memories of the
color and motion characteristics of objects reside in separate locations in the temporal
lobe and thus that the medial temporal region must have multiple sensory inputs.
The macaque monkey has been the principal subject
of anatomical study on the medial temporal region,
and there are likely few differences between macaques
and humans in this respect. Three medial temporal cortical
regions, in addition to the hippocampus and amygdala,
take part in explicit memory. As illustrated in
Figure 13-9, these regions,which lie adjacent to the hippocampus,
are the entorhinal cortex, the parahippocampal
cortex, and the perirhinal cortex (from the Greek
para, meaning “beside,” and rhinal, from the rhinal, or nasal,
sulcus on the bottom of the brain).
A sequential arrangement of connections, charted in
Figure 13-10, shows that the major cortical regions project
to the perirhinal and parahippocampal cortices, which in
turn project to the entorhinal cortex. The prominent input
In Review .
To identify the circuits responsible for memory, it is important to separate explicit memory
from implicit memory. Explicit memory relies on the anterior part of the hippocampus,
the amygdala, and the adjacent cortex. These areas were damaged in H. M.’s brain,
and so he had no explicit memory. An implicit-memory deficit indicates deterioration of
the basal ganglia characteristic of Parkinson’s disease, as seen in patients such as J. K.
496 ! CHAPTER 13
Amygdala
Perirhinal
cortex
Parahippocampal
cortex
Entorhinal
cortex
Hippocampus
Figure 13-9
Medial Temporal Cortex and
Subcortical Structures The brain of a
rhesus monkey viewed ventrally, showing
the medial temporal regions on the left.
Each plays a distinct role in processing
sensory information for memory storage.
The two subcortical structures on the
right, the hippocampus and amygdala,
are not directly visible from the surface
of the brain, because they lie within
the medial temporal cortical regions
illustrated on the left. All these cortical
and subcortical structures are present on
both sides of the brain.
Frontal, parietal, temporal,
occipital, and cingulate cortices
Perirhinal
cortex
Parahippocampal
cortex
Entorhinal
cortex
Hippocampus
Figure 13-10
Reciprocal Medial Temporal Connections
Input from the sensory cortices flows to the
parahippocampal and perirhinal regions, then to the
entorhinal cortex, and, finally, to the hippocampus,
which then feeds back to the medial temporal
regions that connect to the neocortical regions.
CH13.qxd 2/18/05 9:33 AM Page 496
ls
le
ll
from the neocortex to the perirhinal region is from the visual regions of the ventral
stream coursing through the temporal lobe. The perirhinal region is thus a prime candidate
for visual object memory.
Similarly, the parahippocampal cortex has a strong input from regions of the parietal
cortex believed to take part in visuospatial processing. Thus, the parahippocampal
region likely has a role in visuospatial memory. Because both the perirhinal and the
parahippocampal regions project to the entorhinal cortex, this region likely participates
in more-integrative forms of memory. Indeed, the entorhinal cortex is the first to show
cell death in Alzheimer’s disease. Recall from Chapter 5 that Alzheimer’s disease is a
form of dementia characterized by severe deficits in explicit memory (see “Alzheimer’s
Disease” on page 498).
THE HIPPOCAMPUS AND SPATIAL MEMORY
We are left with a conundrum. If the hippocampus is not the key structure
in explicit memory yet is the recipient of the entorhinal connections,
what does it do? The answer is that the hippocampus is probably
engaged in processes requiring the memory for places, such as recalling
the location of an object. This idea was first advanced by John O’Keefe
and Lynn Nadel in 1978.
Certainly, both laboratory animals and human patients with selective
hippocampal injury have severe deficits in various forms of spatial
memory. Rats with hippocampal damage have great difficulty solving
spatial-navigation tasks such as those shown in Figure 13-3. Similarly,
monkeys with hippocampal lesions have difficulty learning the location
of objects, as can be demonstrated in tasks such as those illustrated in
Figure 13-11.
Monkeys are trained to displace objects to obtain a food reward
(Figure 13-11A).After they have learned how to do so, they are given one
of two tasks. In the visual-recognition task shown in Figure 13-11B, the
animal displaces a sample object to obtain a food reward. After a short
delay, the animal is presented with two objects, one of which is novel.
The task is to learn that the novel object must be displaced to obtain a
food reward. This task is a test of explicit visual object memory. Monkeys
with perirhinal lesions are impaired at the visual-recognition task.
In the object-position task in Figure 13-11C, the monkey is shown
one object, which is displaced for a food reward. Then the monkey is
shown the same object along with a second, identical one. The task is to
learn that the object that is in the same position as it was in the initial
presentation must be displaced.Monkeys with hippocampal lesions are
selectively impaired at this object-position task.
From the results of these studies on the hippocampus, we would
predict that animals with especially good spatial memories should have
bigger hippocampi than do species with poorer spatial memories.
David Sherry and his colleagues (1992) tested this hypothesis in birds.
Many birds are cachers: they take foods, such as sunflower seeds, and
hide (cache) them for later consumption. Some birds can find hundreds
of items that they have cached. To evaluate whether the hippocampus
HOW DO WE LEARN AND REMEMBER? ! 497
Entorhinal cortex. Located on the
medial surface of the temporal lobe;
provides a major route for neocortical
input to the hippocampal formation; often
degenerates in Alzheimer’s disease.
Parahippocampal cortex. Cortex
located along the dorsal medial surface of
the temporal lobe.
Perirhinal cortex. Cortex lying next to
the rhinal fissure on the base of the brain.
(B) Visual-recognition task
(C) Object-position task
The monkey is shown one
object to displace for a
food reward.
–
+
– +
+
A monkeys is shown
an object,…
+
…which it then displaces to
obtain a food reward.
A monkeys is trained to
displace an object to
obtain a food reward.
The monkey is then shown
two objects, and the task is
to displace the new object
to obtain the reward.
On the next trial, the
monkey is shown two
identical objects and must
choose the one that is in
the same location as in the
initial presentation.
(A) Basic training
Figure 13-11
Two Memory Tasks for Monkeys (A) In “basic training,” a monkey learns
to displace an object to obtain a food reward. (B) and (C) The plus and minus
signs indicate whether the object is (!) or is not (") associated with food.
CH13.qxd 2/18/05 9:33 AM Page 497
ls
le
ll
plays a role in this activity, Sherry and his coworkers measured hippocampal size in
bird species that are closely related, but only one of which is a food cacher.
As shown in Figure 13-12, the hippocampal formation is larger in birds that cache
food than in birds that do not. In fact, the hippocampus of food-storing birds is more
than twice as large as expected for birds of their brain size and body weight. Sherry
found a similar relation when he compared different species of food-storing rodents.
498 ! CHAPTER 13
Alzheimer’s Disease
Focus on Disorders
That the brain undergoes atrophy with aging was noted in
the 1880s, but the reason was not really understood until
German physician Alois Alzheimer published a landmark
study in 1906. Alzheimer reported on a 51-year-old woman
for whom he described a set of behavioral symptoms and associated
neuropathology. In particular, the woman was demented
and had various abnormalities in the cellular
structure of the cerebral cortex, including both the neocortex
and the limbic cortex.
An estimated 1 million people are now affected by
Alzheimer’s disease in the United States, although the only
certain diagnostic test remains postmortem examination of
cerebral tissue. The disease progresses slowly, and many
people with Alzheimer’s disease probably die from other
causes before the cognitive symptoms become incapacitating.
We knew of a physics professor who continued to work
until he was nearly 80 years old, at which time he succumbed
to a heart attack. Postmortem examination of his
brain revealed significant Alzheimer’s pathology. His slipping
memory had been attributed by his colleagues to “old
timer’s disease.”
The cause of Alzheimer’s disease remains unknown, although
it has been variously attributed to genetic predisposition,
abnormal levels of trace elements (e.g., aluminum),
immune reactions, and slow viruses. Two principal neuronal
changes take place in Alzheimer’s disease:
1. Loss of cholinergic cells in the basal forebrain. One treatment
for Alzheimer’s disease, therefore, is to provide
medication to increase acetylcholine levels in the forebrain.
An example is Cognex, which is the trade name for
tacrine hydrochloride, a cholinergic agonist that appears
to provide temporary relief from the progression of the
disease.
2. Development of neuritic plaques in the cerebral cortex.
A neuritic plaque consists of a central core of homogeneous
protein material (amyloid), surrounded by degenerative
cellular fragments. The cortical plaques, illustrated
here, are not distributed evenly throughout the cortex but
are concentrated especially in the temporal-lobe areas related
to memory. The plaques are often associated with
another abnormality, neurofibrillary tangles, which are
paired helical filaments found in both the cerebral cortex
and the hippocampus.
Cortical neurons begin to deteriorate as the cholinergic loss
and plaques develop. The first cells to die are in the entorhinal
cortex, and significant memory disturbance ensues.
Neuritic plaque, as often found in the cerebral cortices of
Alzheimer patients. The amyloid is the dark spot in the center
of the image, which is surrounded by the residue of
degenerated cells.
Cecil Fox/Science Source/Photo Researchers
Neuritic plaque. Area of incomplete
necrosis (dead tissue) consisting of a
central protein core (amyloid) surrounded
by degenerative cellular fragments; often
seen in the cortex of people with senile
dementias such as Alzheimer’s disease.
CH13.qxd 2/18/05 9:33 AM Page 498
ls
le
ll
Rodents such as Merriam’s kangaroo rat, which stores food in various places throughout
its territory, have larger hippocampi than those of rodents such as the bannertail
kangaroo rat, which stores food only in its burrow.Hippocampal size in both birds and
mammals appears to be related to the cognitive demands of foraging and food storing,
which are highly spatial activities.
One prediction that we might make from the Sherry experiments is that people
who have jobs with high spatial demands might have large hippocampi. Taxi drivers
in London fit this category. To qualify for a cab driver’s license in London, candidates
must pass an extensive examination demonstrating that they know the location
of every street in that huge and ancient city.
Eleanor Maguire and her colleagues (2000), using magnetic resonance imaging
(MRI), found the posterior region of the hippocampus in London taxi drivers to be significantly
larger than in the control subjects. This finding presumably explains why this
select group is able to pass a spatial-memory test that most of us would fail miserably.
RECIPROCAL CONNECTIONS FOR EXPLICIT MEMORY
A key feature of the medial temporal pathway of explicit memory is that it is reciprocal.
That is, the connections from the neocortex run to the entorhinal cortex and then back
to the neocortex (see Figure 13-10). These reciprocal connections have two benefits:
1. Signals that the medial temporal regions send back to the cortical sensory regions
keep the sensory experience alive in the brain. Therefore the neural record of an
experience outlasts the actual experience.
2. The pathway back to the cortex means that the neocortex is kept apprised of information
being processed in the medial temporal regions.We shall see that such
feedback does not happen in the basal ganglia systems taking part in implicit
memory, which may help to explain the unconscious nature of implicit memory.
Although we have focused on the role of the medial temporal regions, other structures
also are important in explicit memory. People with frontal-lobe injuries are not
amnesic like H. M. or J. K., but they do have difficulties with memory for the temporal
order of events. Imagine that you are shown a series of photographs and asked to
remember them. A few minutes later, you are asked whether you recognize two photographs
and, if so, to indicate which one you saw first.
H. M. would not remember the photographs. People with frontal-lobe injuries
would recall seeing the photographs but would have difficulty recalling which one they
had seen most recently. The role of the frontal lobe in explicit memory is clearly more
subtle than that of the medial temporal lobe. But just what is that role?
All the sensory systems in the brain send information to the frontal lobe, as do the
medial temporal regions. This information is not used for direct sensory analysis, and
so it must have some other purpose. In general, the frontal lobe appears to have a role
in many forms of short-term memory.
Joaquin Fuster (e.g., Fuster, Bodner, & Kroger, 2000) studied single-cell activity in
the frontal lobe during short-term-memory tasks. For example, if monkeys are shown
an object that they must remember for a short time before being allowed to make a response,
neurons in the prefrontal cortex will show a sustained firing during the delay.
Consider the tests illustrated in Figure 13-13:
A monkey is shown a light, which is the cue, and then must make a response after
a delay.
In the delayed-response task, the monkey is shown two lights in the choice test and
must choose the one that is in the same location as the cue.
HOW DO WE LEARN AND REMEMBER? ! 499
Relative volumetric ratio of
hippocampus to forebrain
Food-storing Non-food-storing
Chickadee
Common
sparrow
Figure 13-12
Inferring Spatial Memory This graph
relates hippocampal volume to forebrain
volume in 3 food-storing (left) and 10
non-food-storing (right) families of
songbirds. The hippocampi of birds that
cache food, such as the black-capped
chickadee, are about twice as large as
the hippocampi of birds, such as the
sparrow, that do not. Data from “Spatial
Memory and Adaptive Specialization of the
Hippocampus,” by D. F. Sherry, L. F. Jacobs, and
S. J. C. Gaulin, 1992, Trends in Neuroscience, 15,
pp. 298–303.
CH13.qxd 2/18/05 9:33 AM Page 499
ls
le
ll
In the delayed-alternation task, the monkey is again shown two lights in the choice
tests but now must choose the light that is not in the same location as the cue.
Finally, in the delayed-matching-to-sample task, the monkey is shown, say, a red
light, and then, after a delay, is shown a red and a green light. The task is to choose the
red light, irrespective of its new location.
Fuster has found that, in each task, certain cells in the prefrontal cortex will fire
throughout the delay. Animals that have not learned the task show no such cell activity.
Curiously, if a trained animal makes an error, the activity of the cells corresponds
to it: the cells stop responding before the error occurs. In a real sense,
the cells have “forgotten” the cue.
People who have chronically abused alcohol can develop an explicitmemory
disturbance known as Korsakoff’s syndrome. Such people have
severe deficits in explicit memory and, in some cases, in implicit memory
as well. This syndrome is caused by a thiamine (vitamin B1) deficiency
that results from poor nutrition (alcohol abusers often neglect to eat) and
the fact that alcohol inhibits the body’s ability to absorb vitamin B1.
The effect of the B1 deficiency is to produce cell death in the medial
part of the diencephalon (the “between brain”; see Chapter 2), including
the medial thalamus and mammillary bodies of the hypothalamus. In
addition, 80 percent of Korsakoff patients have atrophy (loss of cells) of
the frontal lobes.The memory disturbance is probably so severe inmany
Korsakoff patients because the damage includes not only the frontal lobe
but medial temporal structures as well (see “Korsakoff ’s Syndrome” on
page 502).
Mortimer Mishkin and his colleagues (Mishkin, 1982; Murray,
2000) at the U.S.National Institute of Mental Health proposed a circuit
for explicit memory that incorporates the evidence from both humans
and laboratory animals having injuries to the temporal and frontal
lobes. Figure 13-14 presents a modified version of the Mishkin model
that includes not only the frontal and temporal lobes but also the
medial thalamus, which is implicated in Korsakoff ’s syndrome, and the
activating systems from the basal forebrain, which are implicated in
Alzheimer’s disease:
The sensory neocortical areas send their connections to the medial temporal regions,
which are in turn connected to the medial thalamus and prefrontal cortex.
The basal forebrain structures are hypothesized to play a role in the maintenance
of appropriate levels of activity in the forebrain structures so that they can process
information.
The temporal-lobe structures are hypothesized to be central to the formation of
long-term explicit memories.
The prefrontal cortex is central to the maintenance of temporary (short-term) explicit
memories as well as memory for the recency (chronological order) of explicit
events.
Neural Circuit for Implicit Memories
Hypothesizing that the basal ganglia are central to implicit memory,Mishkin and his colleagues
(Mishkin, 1982;Mishkin et al., 1997) also proposed a circuit for implicit memories.
As Figure 13-15 shows, the basal ganglia receive input from the entire neocortex and
500 ! CHAPTER 13
Time
Cue Delay Choice
Orange juice
reward
In the following tests, a
monkey is shown a
light, which is the cue,
and then it makes a
response after a delay.
Delayed-response task: The
monkey must choose the
light that is in the same
location as the cue.
Delayed-alternation task: The
monkey must choose the
light that is not in the
same location as the cue.
Delayed-matching-to-sample
task: The monkey must
choose the light that is the
same color as the cue.
Off
On
Figure 13-13
Testing Short-Term Memory (Top) A
monkey performing a short-term
memory task responds by pressing the
disc to get a fruit juice reward. (Bottom)
The correct disc varies, depending on the
requirements of the task. The correct
choice for each task is indicated by an
arrow. Adapted from Memory in the Cerebral
Cortex (p. 178), by J. Fuster, 1995, Cambridge,
MA: MIT Press.




Aidan O’Brien saddles century of Royal Ascot winners and shows no sign of
letting up
-
-
Scandinavia beats Trawlerman in compelling Gold Cup
-
‘Very special day for myself and everybody in Ballydoyle’
For all the ...
34 minutes ago
























.jpg)

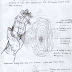

.jpg)
.jpg)

.jpg)



.jpg)




.jpg)

